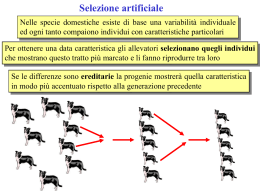
Cap. 22 Genetica di popolazioni, pp. 661-713 Sintesi 22 1. Una popolazione è caratterizzata dalle frequenze dei diversi genotipi e dei diversi alleli al suo interno 2. Una popolazione si dice panmittica o in equilibrio quando le sue frequenze genotipiche possono essere predette sulla base delle frequenze alleliche, e le frequenze alleliche non cambiano attraverso le generazioni 3. I fattori che provocano scostamento dall’equilibrio comprendono unione non casuale dei gameti, mutazione, selezione, migrazione e gli effetti del caso 4. Popolazioni in equilibrio non si evolvono. I fattori che provocano scostamento dall’equilibrio sono i fattori dell’evoluzione Individui, popolazioni Gli individui hanno i loro alleli e genotipi, le popolazioni le loro frequenze alleliche e genotipiche Ogni individuo è come è, le frequenze alleliche cambiano nel tempo e questa è l’evoluzione: genetica evoluzionistica Gli individui si ammalano e guariscono, le popolazioni sono caratterizzate da frequenze di alleli patologici: epidemiologia genetica Gli individui nascono e muoiono, le popolazioni hanno dimensioni fluttuanti e possono estinguersi: genetica della conservazione Prima di tutto: non c’è genetica senza variabilità E la variabilità è prodotta dal processo di mutazione La genetica di popolazioni studia i polimorfismi Polimorfismi morfologici Polimorfismi elettroforesici Polimorfismi di restrizione (RFLP) Polimorfismi di lunghezza Polimorfismi di sequenza Polimorfismi di comportamento Quand’è che una popolazione può dirsi variabile? A N alleli = 5 HO = 0.4 B N alleli = 2 HO = 0.6 Quando il genotipo individuale è difficile da prevedere Elettroforesi L’elettroforesi separa macromolecole in relazione alla loro carica o alle loro dimensioni Frequenze alleliche F S S FS S FF SS SS FS SS FS F FS FS S fenotipo osservato FS Freq. genotipiche: FF=0,2 FF FS FS=0,4 Freq. alleliche: f(F)=p, f(S)=q p = (NF + ½ NH) / NT p = (2 + 2)/10 = 0,4 q = (4 + 2)/10 = 0,6 FS SS genotipo dedotto SS=0,4 p+q=1 = (2NF + NH)/2NT p= (4 + 4)/20 = 0,4 q = (8 + 4)/20 = 0,6 p+q = 0,4 + 0,6 = 1 Che relazione lega frequenze alleliche e genotipiche? Generazione 0: N(AA) = d N(Aa) = h N(aa) = r Totale = T F(A) = (NF + ½ NH) / NT = (d + ½ h) / T = p Assunzione: l’unione fra i genotipi è casuale. Panmissia Conseguenza: l’unione fra i gameti è casuale Perciò alla generazione 1 avremo: F(AA) = p2 F(Aa) = 2pq F(aa) = q2 Totale =1 1. Se l’unione dei gameti è casuale, le frequenze genotipiche dipendono dalle frequenze alleliche secondo la relazione Fr. alleliche: p, q Fr. Genotipiche: p2, 2pq, q2 Frequenze alleliche alla generazione 1: F(A) = (NF + ½ NH) / NT = (p + ½ 2 pq) / 1 = p 2. Se l’unione dei gameti è casuale, le frequenze alleliche non cambiano attraverso le generazioni: Equilibrio di Hardy-Weinberg Accoppiamento casuale o random mating MATING AA x AA (p2)(p2) AA x Aa (p2)(2pq) AA x aa (p2)(q2) Aa x AA (2pq)(p2) Aa x Aa (2pq)(2pq) Aa x aa (2pq)(q2) aa x AA (q2)(p2) aa x Aa (q2)(2pq) aa x aa (q2)(q2) MAT. FREQ. PROGENIE Aa p4 AA p4 2p3q p3q p3q p2q2 aa p2q2 2p3q p3q p3q 4p2q2 p2q2 2p2q2 p2q2 2pq3 pq3 pq3 p2q2 p2q2 2pq3 pq3 q4 pq3 q4 E alla fine nella progenie f(AA) = p4 + 2p3q + p2q2= p2 (p2+ 2pq +q2) = p2 f(Aa) = 2p3q + 4p2q2 + 2pq3 = 2pq (p2 + 2pq +q2) = 2pq f(aa) = p2q2 + 2pq3 + q4 = q2 (p2 + 2pq +q2) = q2 Cioè esattamente le frequenze che si ottengono immaginando di accoppiare a caso i gameti del pool genico parentale Dopo la prima generazione di panmissia p, q p2, 2pq, q2 Quindi, in una popolazione panmittica: •Le frequenze genotipiche dipendono esclusivamente dalle frequenze alleliche della generazione precedente •Le frequenze alleliche non cambiano attraverso le generazioni Quindi, se c’è equilibrio non c’è evoluzione, e viceversa L’equilibrio di Hardy-Weinberg Dopo una generazione di accoppiamento casuale: Genotipo AA Aa aa Frequenza p2 2pq q2 Cosa vuol dire equilibrio Se in una popolazione ho 80 alleli a e 120 A: Allora p = 80/200 = 0,4, q = 0,6 0,16, 0,48, 0,36 Possibili distribuzioni dei genotipi: AA Aa aa 40 35 20 16 10 0 0 10 40 48 60 80 60 55 40 36 30 20 p 0,4 0,4 0,4 0,4 0,4 0,4 Condizioni per l’equilibrio di Hardy-Weinberg • • • • • • • • Organismo diploide, riproduzione sessuata Generazioni non sovrapposte Unione casuale Popolazione grande Mutazione trascurabile Migrazione trascurabile Mortalità indipendente dal genotipo Fertilità indipendente dal genotipo Se non si incontrano queste condizioni: • • • • • • Unione casuale Inbreeding Popolazione grande Deriva genetica Mutazione trascurabile Mutazione Migrazione trascurabile Migrazione Mortalità indipendente dal genotipo Selezione Fertilità indipendente dal genotipo Selezione Studio di polimorfismi di inserzione al locus PLAT Frequenze genotipiche ed alleliche: Inserzioni al locus PLAT in 27 popolazioni umane Deriva genetica significa che c’è una componente casuale nel successo riproduttivo Riproduzione asessuata; N costante; ogni individuo lascia 1 discendente Riproduzione asessuata; N costante; numero variabile di discendenti Esperimento di Buri (1956) 109 popolazioni 8 maschi, 8 femmine Inizialmente, tutti eterozigoti bw+ bw Condizioni dell’esperimento di Buri • • • • • • • • Organismo diploide, riproduzione sessuata Generazioni non sovrapposte Unione casuale Popolazione grande Mutazione trascurabile Migrazione trascurabile Mortalità indipendente dal genotipo Fertilità indipendente dal genotipo Simulazione di deriva genetica in popolazioni diploidi di 10000 e 4 individui La deriva riduce la variabilità entro popolazioni e aumenta quella fra popolazioni Perché è importante la deriva genetica? • Importanza evolutiva: cambiamento non adattativo, specie in piccole popolazioni • Importanza per la conservazione: perdita di diversità genetica, specie in piccole popolazioni • Importanza biomedica: alleli patologici altrove rari possono essere comuni in piccole popolazioni Colli di bottiglia Variabilità genetica nel ghepardo Acinonyx Jubatus Jubatus (S. Africa) 2,500 (Namibia) 1,500 (Botswana) 1,500 (Kenya/Tanzania) Acinonyx Jubatus Rainey (E. Africa) less than 1,000 Acinonyx Jubatus Hecki (N. Africa) less than 1,000 Acinonyx Jubatus Venaticus (Asia) virtually extinct Acinonyx Jubatus Raddei (Iran/Turkestan) approx. 200 Livelli di eterozigosi per marcatori VNTR N H media A. jubatus jubatus 7 0.280 A. jubatus raineyi 9 0.224 Felis catus 17 0.460 Panthera Leo (Serengeti) 76 0.481 Panthera Leo (Ngorongoro) 6 0.435 Menotti-Raymond & O’Brien 1993 Bottleneck datato al Pleistocene Flusso genico Migrazione di individui (dalla popolazione A alla popolazione B) Colonizzazione di habitat precedentemente non occupati Dispersione dei propaguli Flusso genico e deriva hanno effetti opposti Il flusso genico introduce nuovi alleli nelle sottopopolazioni e riduce le differenze fra sottopopolazioni Flusso genico deriva Nota bene La variabilità interna di una popolazione è solo uno degli aspetti della variabilità genetica: Variabilità tra individui della stessa popolazione Variabilità tra individui di popolazioni diverse Variabilità tra individui di gruppi di popolazioni diverse eccetera Selezione e variazione delle frequenze alleliche: melanismo Frequenza della forma melanica (DD e Dd) in Nord America (Grant & Wiseman 2002 ) Per l’evoluzione darwiniana servono quattro cose 1. Esiste una variabilità 2. Questa variabilità è, almeno in parte, ereditaria 3. Nel contesto ambientale in cui si trovano, individui con caratteristiche ereditarie diverse hanno fertilità o mortalità diverse 4. Le caratteristiche degli individui che lasciano più discendenti si diffondono nella popolazione: adattamento Evoluzione darwiniana 1. Esiste una variabilità Evoluzione darwiniana 2. Questa variabilità è, almeno in parte, ereditaria Evoluzione darwiniana 3. Nel contesto ambientale in cui si trovano, individui diversi hanno fertilità o mortalità diverse Verdi = 5/15 = 33% Evoluzione darwiniana 3. Nel contesto ambientale in cui si trovano, individui diversi hanno fertilità o mortalità diverse Evoluzione darwiniana 4. Le caratteristiche degli individui che lasciano più discendenti si diffondono nella popolazione: adattamento Verdi = 4/8 = 50% Ma non dimentichiamo il caso (1) Verdi = 0% Ma non dimentichiamo il caso (2) Crescita demografica Verdi = 7% Fitness Misura la speranza di progenie. Ha due componenti, legate a differenze in fertilità e mortalità Genotipo AA Aa aa N medio discendenti 2.4 2.4 3.0 Fitness riproduttiva 2.4/3.0 =0.8 2.4/3.0=0.8 3.0/3.0=1.0 Genotipo AA Aa aa Vita riproduttiva media 2.0 1.8 1.4 1.8/2.0 =0.9 1.4/2.0 =0.7 Fitness di sopravvivenza 2.0/2.0 =1.0 Fitness Complessivamente: Genotipo AA Aa aa Fitness riprod. 0.8 0.8 1.0 Fitness sopravv. 1.0 0.9 0.7 Prodotto 0.8 0.72 0.7 Fitness totale ω 0.8/0.8=1.0 0.72/0.8=0.9 0.7/0.88 Coefficiente di selezione: s = 1 – ω Misure sperimentali della fitness in Biston betularia Birmingham Dorset Genotipo DD + Dd dd DD + Dd dd Rilasciate 154 64 406 393 Ricatturate 82 16 19 54 0.53 0.25 0.047 0.137 1 0.47 0.343 1 Sopravvivenza ω Anemia falciforme Correlazione fra la frequenza di HbS e la presenza di malaria endemica wHb/Hb = 0.88; wHb/HbS = 1.0 wHbS/HbS = 0.14 Stima empirica dei coefficienti di selezione per HbS in Nigeria Ma perché, dopo tutta questa selezione, non siamo diventati perfetti? 1. L’ambiente cambia “Stone agers in the fast lane” “Red queen” hypothesis (John Maynard Smith) 2. Selezione sessuale In specie sessualmente dimorfiche le femmine scelgono il partner in base a criteri che possono promuovere o meno l’adattamento. Due ipotesi principali: 1. “Good genes” 2. “Sexy sons” Riassunto •Si possono predire le frequenze genotipiche in una popolazione panmittica •La deriva genetica porta le popolazioni a diventare internamente più omogenee e a differenziarsi fra loro •Il flusso genico porta le popolazioni a diventare più omogenee fra loro e a differenziarsi internamente •La selezione naturale fa aumentare le frequenze di alleli associati a maggiore fertilità o minore mortalità •L’evoluzione è il risultato dell’interazione fra questi fattori sulla variabilità generata dal processo di mutazione
Scaricare