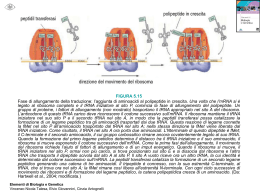
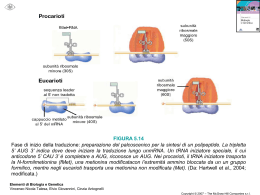
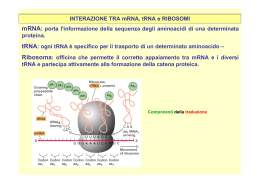
Biologia Molecolare CDLM in CTF 2010-2011 – La modificazione dell’RNA e la traduzione La maturazione del trascritto primario I microRNA Le componenti del macchinario di traduzione Il meccanismo della traduzione La maturazione dell’RNA messaggero procede attraverso vari passaggi: Rimozione degli introni e giunzione degli esoni (SPLICING) Aggiunta di una metil-guanosina al 5’ Aggiunta di una coda di poliA al 3’ Lo splicing è stato osservato in organismi semplici come un processo indipendente dall’intervento dello spliceosoma Lo spliceosoma interviene nella maturazione della maggior parte degli mRNA degli eucarioti pluricellulari 5 piccoli RNA (snRNA) Più di 200 proteine I complessi si chiamano snRNP (U1, U2, U4, U5 e U6) Fare scansione • Resistenza alle 5’deossiribonucleasi • Efficienza dello splicing • Riconoscimento dell’estremità per la traduzione Poli(A) Polimerasi La maturazione del trascritto primario I microRNA Le componenti del macchinario di traduzione Il meccanismo della traduzione I siRNA sono piccoli RNA che operano autosilenziamento (stesso locus) Sono di solito complementari al 100% alle sequenze bersaglio I microRNA (o miRNA) sono coinvolti nei processi di eterosilenziamento La complementarietà è inferiore al 100% I trascritti primari dei geni miRNA sono chiamati primiRNA. I pri-miRNA vengono tagliati da un enzima chiamato Drosha in molecole più piccole, a doppio filamento, chiamate premiRNA. • I pre-miRNA vengono esportati nel citoplasma e tagliati in RNA doppio filamento più piccoli da un altro enzima chiamato Dicer. • Uno dei due filamenti contiene il miRNA maturo, lungo solitamente tra i 19 e i 25 nucleotidi, che viene incorporato in un complesso proteico chiamato RISC. I miRNA nei RISC sono in grado di legarsi a siti specifici di mRNA provocandone il silenziamento: L’appaiamento della sequenza del miRNA con il suo sito bersaglio non è perfetto, ma può contenere bulge e loop. Dalle coppie miRNA/target individuate sperimentalmente emergono alcune regolarità nelle modalità di appaiamento. La regione iniziale (5’) del miRNA è chiamata seed e sembra essere la regione più importante nel silenziamento. E’ lunga solitamente tra i 7 e i 10 nucleotidi, ma esistono casi di seed più corti o più lunghi. Tale regione è solitamente appaiata in modo perfettamente complementare al suo target: Il primo nucleotide del miRNA non determinante e può non essere appaiato. è • La regione a valle del seed contiene solitamente un bulge o un loop: • La regione 3’ del miRNA mostra solitamente una complementarità imperfetta al suo target. • Le coppie G:U nella regione del seed sembrano essere sfavorevoli al silenziamento, sebbene siano ammesse, mentre sono abbastanza comuni nella regione 3’. • Infine, le regioni di legame dei miRNA si trovano nella regione 3’ UTR degli mRNA bersaglio. I miRNA svolgono un ruolo centrale nel controllo di numerosi processi fisiologici: Aberrazioni nella loro espressione (mancanza, sotto o sovra espressione) sono correlate a diversi tipi di patologie: Sviluppo Differenziamento cellulare Apoptosi Cancro Malattie neurodegenerative Malattie cardiache Si tratta dunque di molecole molto importanti, il cui studio è fondamentale nella comprensione dei processi biologici, dei fenotipi patologici e, di conseguenza, nel design di terapie innovative. La maturazione del trascritto primario I microRNA Le componenti del macchinario di traduzione Il meccanismo della traduzione Procarioti: ribosomi 70s 50s grande 30s piccola Eucarioti: ribosomi 80s 60s grande 40s piccola Sono molecole adattatrici che legano e trasportano aminoacidi specifici. Due porzioni particolarmente importanti sono: • ANTICODONE : sequenza di 3 nucleotidi complementari al codone corrispondente all’aminoacido legato al tRNA. • Sito di attacco dell’aa. : sequenza 5’CCA-3’, uguale in tutti tRNA. MET L’anticodone si appaia in maniera specifica al codone sull’mRNA in modo da permettere al tRNA di trasportare l’aminoacido corrispondente quando questo viene richiesto dalla informazione contenuta sul messaggero MET AUG UUAG CUUC GCGA UAUC Quanti sono i tRNA? UNA LOGICA DEDUZIONE PER QUANTO DETTO FIN’ORA E’ CHE DOVREBBERO ESSERCI 61 tRNA DIVERSI, UNO PER CIASCUN CODONE; CIO’ IMPLICHEREB BE DUE COSE: 1. GRANDE CONSUMO ENERGETICO DA PARTE DELLA CELLULA; 2. tRNA AVENTI ANTICODONI DIFFERENTI DEVONO PORTARE LO STESSO aa. IN REALTA’ QUESTA CONDIZIONE E’ VERA SOLO PER ALCUNI tRNA, PER LA MAGGIORPARTE (NELL’UOMO ESISTONO INFATTI SOLO 48 ANTICODONI DIFFERENTI A FRONTE DEI TEORICI 61 POSSIBILI), PERO’, SUSSISTE IL FENOMENO DEL “VACILLAMENTO” DELLA TERZA BASE, PER CUI SI POSSONO FORMARE LEGAMI ANCHE “NON ORTODOSSI” FRA IL NUCLEOTIDE IN POSIZIONE 5’ DEL tRNA E IL NUCLEOTIDE IN 3’ DELL’mRNA. • Adenilazione aminoacido • AMP rilasciato e legame al tRNA • Rilascio aminoacil tRNA La maturazione del trascritto primario I microRNA Le componenti del macchinario di traduzione Il meccanismo della traduzione Le fasi della traduzione nei procarioti 1) Inizio: la subunità minore del ribosoma si lega al messaggero. Il primo tRNA (quello della fMET) con anticodone complementare alla tripletta AUG, che rappresenta il segnale di inizio, si lega in corrispondeza del codone. Questo scatena il legame della subunità maggiore e l’inzio della trascrizione. 2) Allungamento: il ribosoma si sposta in direzone 3’ sul mRNA. I tRNA si lagano uno dopo l’altro ai codoni che via via vengono incontrati dal ribosoma mentre “legge” il filamento ribonucleotidico. Il ribosoma catalizza di volta in volta il legame tra l’aminoacido già presente all’interno del sito attivo ed il nuovo aminoacido appena entrato nel ribosoma allungando la catena proteica nascente di un aminoacido alla volta. 3) Terminazione: Quando il ribosoma incontra un segnale di stop nessun tRNA si lega al sito attivo del ribosoma. Si lagano invece specifici fattori di terminazione proteici che interrompono la traduzione e facilitano la destabilizzazione del complesso. La Fase di inizio nei procarioti 1) Le subunità minori libere (non legate a subuinità maggiori) e legate al fattore proteico di inizio IF-3 sono ingrado di legarsi all’RNA messaggero in corrispondenza di una regione particolare contenente: • Un esanucleotide chiamato sequenza di Shine-Dalgarno (AGGAGG) complementare ad un tratto dell’RNA ribosomiale 16s • Un codone di start AUG a circa 10 basi dalla sequenza Shine-Dalgarno 2) Il fattore proteico di inizio IF-2 si lega in corrispondenza di un sito particolare del ribosoma, il sito P che è ancora nel suo stato parziale perché è costituito soltanto dalla subunità minore. Il legame di IF-2 permette l’attacco del primo aminoacil-tRNA, quello della formil metionina chiamato fMettRNAf 3) Questo complesso permette alla subuinità maggiore di legare la subunità minore già posizionata sull’mRNA. Il complesso viene facilitato da un altro fattore proteico chiamato IF-1 4) L’associazione delle due subunità provoca il rilascio di tutti i fattori proteici di inizio (IF) Soltanto il fMet-tRNAf è capace di legarsi al sito P parziale ed è questo il motivo perché il primo codone AUG viene riconoscuto da questo aminoacil-tRNA mentre gli AUG interni vengono riconosciuti dal Met-tRNA m L’inizio della traduzione nei procarioti fMet Large subunit E P A La traduzione inizia col riconoscimento dell’mRNA da parte della subunità minore. Il legame del tRNA della fMET (con anticodone complementare ad AUG) permette il legame della subunità maggiore. UAC 5’ GAG...CU-AUG--UUC--CUU--AGU--GGU--AGA--GCU--GUA--UGA-AT GCA...TAAAAAA Small mRNA subunit 3’ La Fase di allungamento nei procarioti 1) All’inizio della fase di allungamento il sito P si trova occupato dalla formil-metionina legata al suo tRNA, il sito A è disponibile per l’interazione di un nuovo aminoacil-tRNA a patto che l’anticodone sia complementare al codone successivo all’AUG 2) L’aminoacil-tRNA entra nel sito A soltanto se legato ad un fattore di allungamento: EF-Tu. Questo fattore deve trovarsi nella sua forma “carica” cioè legato a GTP. 3) Una volta dentro il sito A, la subuinità maggiore del ribosoma catalizza l’idrolisi del GTP in GDP il che fornisce l’energia per creare il nuovo legame peptidico tra il nuovo aminoacido a la fMet. 4) L’EF-Tu scarico (col GDP) si stacca e va a finire nel citoplasma dove viene RICARICATO da un altro fattore di allungamento (EF-TS), tornando ad essere disponibile per il legame ad un altro aminoacil-tRNA 5) Nel frattempo il ribosoma effettua la TRASLOCAZIONE. Scorre sull’RNA messaggero verso la direzione 3’ spostando il tRNA iniziatore nel sito E e il tRNA legato al peptide nascente nel sito P. Il sito A è di nuovo libero per la interazione di un nuovo aminoacil-tRNA 6) La fase di allungamento si itera dal punto 1 La fase di allungamento Un nuovo aminoacil tRNA (complesso tra tRNA ed il suo aminoacido corrispondente) entra nel sito A del ribosoma. Il suo anticodone è complementare al codone che si trova in corrispondenza del sito A. Il primo aminoacido (la metionina) presente nel sito P si troverà quindi vicino ad un nuovo aminoacido appena trasportato dal suo tRNA. La subunità maggiore del ribosoma catalizza il legame di questo nuovo aminoacodo con la metionina. Il legame della metionina con il suo tRNA viene rotto. La fase di allungamento A questo punto abbiamo un tRNA “scarico” nel sito P, un tRNA legato alla catena nascente (di due aminoacidi, al momento) nel sito A. Il ribosoma si sposta a sinistra sull’mRNA: il tRNA vuoto viene liberato nel citoplasma, il tRNA legato a 2 aminoacidi viene spostato nel sito P ed il sito A è di nuovo disponobile per l’attacco di un nuovo aminoacil-tRNA. Il processo viene iterato allungando ogni volta la catena nascente di un aminoacido, seguendo le istruzioni scritte sul messaggero. La fine della traduzione Quando il ribosoma, legato alla catena proteica nascente, raggiunge un codone di stop, invece di un aminoaciltRNA, nel sito A si lega un fattore di terminazione. La proteina nascente lascia il suo tRNA, il ribosoma si destabilizza e la sintesi della proteina finisce. La traduzione negli eucarioti La traduzione procede in maniera abbastanza simile negli eucarioti anche se ci sono alcune piccole differenze. Il tRNA iniziatore per la metionina, sebbene sia strutturalmente e funzionalmente diverso da quello utilizzato nei codoni AUG interni trasporta una metionina non modificata. I fattori accessori sono più numerosi Il riconoscimento della regione di attacco della subunità minore non è mediato dalla sequenza di Shine-Dalgarno ma dalla presenza del cap il 5’. Una serie di proteine formano dei complessi che interagendo permettono al ribosoma di posizionarsi in maniera corretta.
Scaricare