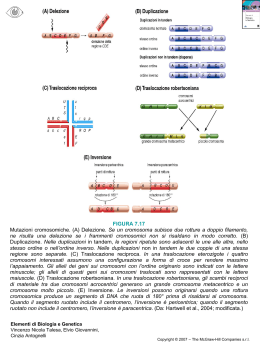
LE MUTAZIONI Buongiorno a tutti, io sono una collaboratrice del professor Fazio e oggi lo sostituisco perché non può venire. Questa lezione verte soprattutto sulle mutazioni, meccanismi di formazioni di mutazioni, classificazioni e patologie ad esse associate. Due punti principali quando si parla delle mutazioni sono i meccanismi di formazione delle mutazioni e le diverse categorie in cui le mutazioni possono essere classificate. Inizialmente una suddivisione riguarda le mutazioni germinali e somatiche. Le prime riguardano la linea germinale e sono quelle ereditabili, vengono cioè trasmesse dai genitori alla prole. Le mutazioni possono essere geniche, cromosomiche e gnomiche. Le geniche interessano il gene o parte di esso, le cromosomiche interessano una parte di cromosoma, o un intero cromosoma o più cromosomi, le genomiche coinvolgono gran parte del genoma e spesso sono incompatibili con la vita e sono le più frequenti nei tumori. Il professore vi avrà sicuramente parlato di polimorfismi e questo è un collegamento da fare con le mutazioni perché non tutti i difetti informazionali determinano una alterazione fenotipica identificabile perché anche una mutazione può dare origine ad un polimorfismo che andrà a determinare quella variabilità genetica tipica della vita. Le mutazioni possono coinvolgere sia la sequenza del DNA ma possono essere dovute anche a fattori ambientali,come fattori fisici, chimici, biologici e intrinseci. Tra i fattori fisici abbiamo le radiazioni che possono essere naturali,da inquinamento o che derivano da trattamenti terapeutici come i raggi x, e da qui la osservazione di non sottoporsi frequentemente ai raggi x per il loro effetto mutageno sul DNA da overesposizione. I fattori chimici sono forti fattori mutageni, e possono essere ambientali endogeni, da inquinamento, e farmaci. Ci sono poi i fattori biologici come i virus. MUTAZIONI GENICHE Tra le mutazioni geniche le più caratteristiche sono le puntiformi che interessano singole basi o dei tratti costituiti da quelle basi. Si suddividono in: sostituzioni di una base con un’altra e possono essere delle transizioni se riguardano sostituzione di una purina con un’altra purina o transversione se riguardano la sostituzione di una purina con una pirimidina; inserzioni se viene inserita una base nel filamento nascente e quindi in questo caso avremo un filamento con un numero maggiore di basi rispetto al filamento parentale; delezione è la sottrazione di una base duplicazione è la duplicazione di due o più basi; l’inversione è quando una coppia di basi viene ruotata di 180°. Ovviamente gli effetti funzionali che le mutazioni puntiformi hanno sul prodotto genico variano a seconda delle sequenze in cui queste mutazioni si trovano, quindi a seconda che si trovano su delle sequenze codificanti o su delle sequenze regolative avremo degli effetti differenti. Per quanto riguarda le sequenze codificanti, le sostituzioni possono portare a delle mutazioni che si dividono in senso, non senso e dissenso a seconda che la mutazione fenotipica porti ad un cambiamento dell’amminoacido corrispondente oppure se porta alla formazione di un codone di stop, oppure ci possono essere delle mutazioni senso in cui la mutazione porta alla produzione dello stesso amminoacido. Ovviamente l’effetto che queste sostituzioni hanno sulla proteina è un effetto funzionale sul prodotto perché la proteina molte volte può esser troncata o pure può essere mutata e non svolgere più la propria funzione. Le inserzioni e delezioni causano ovviamente uno slittamento della lettura del DNA, un “frame shift”. Le mutazioni puntiformi possono anche avvenire sulle sequenze regolative, quindi sugli splicing, sugli introni, www.slidetube.it Pagina 1 sulle sequenze enhancer del promotore e in questo caso avremo delle mutazioni sia in mRNA, sia sull’espressione della proteina, sia sulla funzionalità della proteina. Per quanto riguarda invece i meccanismi di formazione delle mutazioni, sono essenzialmente due: l’errore di duplicazione e l’effetto del mutageno. Per quanto riguarda l’errore di replicazione, al momento si ha una molecola parentale normale, quindi ci cono i due filamenti normali, con le basi appaiate in modo corretto. Al momento della replicazione i due filamenti si dividono. Nel primo avremo una duplicazione corretta e quindi il filamento nascente avrà la sequenza complementare alla catena parentale; nel secondo filamento invece si inserisce un errore di replicazione, quindi in questo caso avremo un nucleotide sbagliato che non si appaia correttamente con il nucleotide della sequenza parentale. Ora se non interviene un sistema di riparazione del DNA, questo nucleotide sbagliato rimarrà anche nella seconda onda di replicazione. Quindi nel secondo ciclo di replicazione i due filamenti nuovamente si aprono e la catena nascente presenterà entrambi i filamenti mutati poiché a livello della mutazione la base non si appaia, mentre il filamento normale darà origine ad una molecola uguale a quella normale iniziale. Quindi una sola molecola figlia su 4 presenta la mutazione, quindi se non interviene il meccanismo di riparazione del DNA, dopo due cicli di replicazione la mutazione si fissa e questo è importante da ricordare. Un altro meccanismo che dà origine a mutazioni è l’effetto di un mutageno. In questo caso il mutageno è di natura chimica e si va a posizionare a livello di una base di uno dei filamenti della molecola parentale mascherando il nucleotide e formando una sorta di addotto. Questo addotto fa si che il macchinario di riparazione del DNA non possa intervenie in quanto l’ingombro sterico dovuto all’addotto fa si che la polimerasi non riconosca quello che è il nucleotide originale, cioè quello parentale. Quello che si ha è che al momento della replicazione i due filamenti si aprono, e avremo un figlio normale derivato dal filamento originale non mutato, ed una molecola figlia che invece è mutata perché durante la replicazione la polimerasi in corrispondenza dell’addotto, indicato con la x, mette una base a caso tra le quattro. Si avrà il 25% di probabilità che la base messa caso sia quella corretta, e il 75% che sia quella sbagliata e come abbiamo detto a questo livello no può intervenire la polimerasi proprio perché non sa quale sia la base originale. Nel secondo ciclo di replicazione quello che si ha è che i due filamenti si separano, entrambi i filamenti presentano la mutazione perché uno ha l’addotto, l’altro ha una base che nel 75% dei casi è quella scorretta e quindi avremo la formazione di due molecole entrambe mutate. Per l’effetto dell’addotto avremo quindi due molecole mutate su quattro molecole figlie. Anche qui la mutazione è fissata dopo due cicli di replicazione, perché a livello dei due cicli si pensa che possa intervenire il sistema di riparazione del DNA anche se nel secondo caso e in questo caso è molto più difficile che possa intervenire rispetto all’errore che è avvenuto prima. Questo meccanismo di formazione indotto dal mutageno verrà ripreso dal professore quando parlerà di cangerogenesi chimica, quindi questo lucido è da tenere presente sia per quanto riguarda la cangerogenesi chimica, sia per la fissazione della mutazione nel secondo ciclo di replicazione. Questo è un breve riassunto di quelle che sono le mutazioni puntiformi e quindi si ha la sequenza di DNA originale, la sostituzione per transizione, la sostituzione per transversione, l’inserzione di una coppia di nucleotidi, la delezione, la duplicazione e l’inversione. Un altro meccanismo di formazione delle mutazioni è dovuto alla conformazione delle basi stesse, in quanto si è visto che l’adenina, nella sua conformazione imminica può avere un accoppiamento illegittimo con la citosina e questo provoca una mutazione a livello della sequenza del DNA. Oltre all’accoppiamento illegittimo con la timina, l’adeninina imminica può formare dei dimeri con un’altra base, sempre con la adenina che in questo caso però ruota di 180°. Quindi sono tutti accoppiamenti illegittimi che causano mutazioni. Un’altra causa di mutazione nel DNA è la presenza nel genoma di sequenze ripetute. In questo caso le zone ondeggiate presentano delle sequenze ripetute che possono essere anche distanti tra loro lungo il gene. In questo caso durante la replicazione si ha che la prima parte della sequenza ripetuta viene replicata dalla sequenza nascente. La sequenza nascente si blocca alla fine della prima sequenza ripetuta. La seconda sequenza ripetuta attraverso il 5’ fa una rotazione di 180° e si va ad appaiare con la sequenza di omologia presente sulla sequenza nascente, creando una sorta di loop che comprende la regione della sequenza ripetuta iniziale 1 con la sequenza che è compresa con queste due regioni ripetute. Allora ripeto, ci sono due www.slidetube.it Pagina 2 sequenze ripetute che sono distanziate. Nel momento della replicazione viene sintetizzata la catena nascente che si ferma in corrispondenza della fine della prima sequenza ripetuta. La sequenza due presenta omologia per questa sequenza e quindi può appaiarsi alla complementare sequenza nascente, causando un loop. Queste non sono mutazioni molto frequenti però avvengono. Il loop nel secondo ciclo di replicazione verrà perso quindi avviene una delezione perché non viene riconosciuto dalla polimerasi. Altre mutazioni che si possono avere sono in sequenze in cui ci cono delle basi che sono altamente ripetute, qui ad esempio abbiamo una regione che presenta 5 timine. In questo caso puo’ accadere che una delle timine venga esclusa per abbinamento incorretto e quindi avremo la formazione di filamento parentale che manca di un’adenina e che quindi è diverso dal filamento parentale. Un’altra alterazione che può avvenire è l’inserzione invece, in queste sequenze ripetute, di un’altra base corrispondente sempre alla base presente, quindi avremo nel filamento figlio non più 5 adenine ma 6. Questi sono tutti eventi casuali che possono avvenire. Un’altra tipologia di sequenze ripetute all’interno del genoma è la presenza di triplette che possono essere ripetute e presenti in cromosomi in numero diverso. Nella replicazione normale le tre triplette vengono tutte e tre mantenute; può succedere nel secondo caso che una delle tre triplette venga inserita sempre per errore della polimerasi, quindi in questo caso nel filamento figlio non avremo più tre triplette ma 4 a causa dell’inserzione, oppure che durante la replicazione una delle triplette venga persa, e anche qui avremo delle mutazioni. Malattie tipiche da triplette sono la sindrome dell’X fragile e la fibrosi cistica. Altre cause di mutazioni sono la presenza di pseudogeni (=geni identici al gene funzionale da cui differiscono per la mutazione di una sola base). Durante la replicazione questi pseudogeni possono essere introdotti e fanno sì che la proteina prodotta non sia più funzionale. Vengono riconosciuti e scambiati per il gene normale proprio perchè differiscono per solo una base e causano mutazioni. Un’altra causa di mutazione è la presenza di crossig-over durante la meiosi: invece di avvenir su delle sequenze omologhe, avviene su delle sequenze simili che molte volte sono anche distanti tra loro lungo il gene. In questo caso specifico il crossino-over può dare origina a duplicazioni, quindi nello stesso filamento abbiamo la presenza dello stesso gene due volte. Un altro effetto del crossing-over è la produzione di proteine di fusione nel caso in cui il crossino-over avvenga su due cromosomi diversi. In questo caso si avrà una proteina di fusione in quanto in questo gene ci sarà la proteina che deriva dal cromosoma uno e quella che invece si trova sul cromosoma 2. Questa proteina di fusione è una proteina che molte volte determina dei tumori. Questo è il caso per esempio della leucemia mieloide cronica (bcr able in questo caso). Allora questo è uno schemino che fa vedere come all’interno del genoma sono presenti dei motivi, in particolar modo delle triplette, ripetuti, e come questa situazione sia una situazione normale, in quanto su ogni cromosoma, in percentuale diverse, queste triplette possono essere presenti e non causare mutazioni. In particolare, il numero di queste ripetizioni si aggira intorno a 19-30 o anche 40 per particolari tipi di cromosomi. Per quanto riguarda ad esempio la sindrome dell’x fragile, si ha un numero elevatissimo di triplette che invece di essere intorno a 50 arrivano a 250-400 copie. Passiamo adesso alle MUTAZIONI CROMOSOMICHE. Esse si possono suddividere in quantitative e qualitative a seconda che coinvolgano il numero dei cromosomi o la struttura dei cromosomi. Nelle mutazioni cromosomiche quantitative, ovviamente c’è un numero totale scorretto di cromosomi e si possono avere la poliploidia, l’anaploidia e la mixoploidia. Nella poliploidia abbiamo delle coppie extra di tutti i cromosomi, quindi l’individui invece di avere un corredo normale di 2n, ne avrà uno di 3n nel caso della triploidia, 4n nel caso della tetraploidia, … Nella anaploidia l’individuo avrà una acquisizione o una perdita di soltanto alcuni cromosomi, questo è il caso ad esempio della trisomia 21 in cui il cromosoma 21 è presente in triplice coppia o della monosemia dell’ X (sindrome di Turner) in cui il soggetto fenotipicamente femmina, ha un solo cromosoma X; www.slidetube.it Pagina 3 La mixoploidia è l’unione di due o più linee cellulari che possono derivare da un singolo zigote nel caso del mosaicismo o da più zigoti nel caso della chimera. Ltro lucido Per quanto riguarda la poliploidia vengono qui riportati degli esempi di triploidia e tetraploidia. La triploidia diciamo può essere generata da tre eventi fondamentali: il primo quando la cellula uovo viene fecondata da due spermatozoi e quindi l’individuo avrà corredo cromosomico 3n, dal momento che ogni gamete ha corredo n; il secondo è quando il gamete femminile è 2n; il terzo quando lo spermatozoo anziché essere n è 2n. Nella tetraploidia abbiamo uno zigote normale 2n che nel momento della replicazione avviene solo la replicazione dell’DNA, senza la divisione cellulare. Questi sono invece gli esempi della mixoploidia che si divide in mosaicismo o chimerismo. A livello del mosaicismo abbiamo la formazione di uno zigote a livello precoce dell’embriogenesi, quindi a livello della blastocisti di solito, in cui una delle quattro cellule è interessata da una mutazione, alterazione genica. In seguito alla replicazione avviene che solo la progenie che deriva dalla cellula alterata sarà alterata, mentre la progenie che deriva dalle altre cellule sarà normale. Quindi alla fine si avrà un insieme di foglietti di cui alcuni normali (quelli che derivano dalle cellule normali), alcuni alterati (quelli che derivano dalla cellula anomala). La popolazione eterogenea che si viene a formare presenterà per ¾, dei foglietti normali perché derivano dalle 3 cellule sane, per ¼ dei foglietti alterati perché derivano dalla cellula mutata. Individuo detto a mosaico proprio perché presenta alcuni foglietti alterati, altri no. Nel caso della chimera abbiamo invece due zigoti, d cui uno normale, ed uno che presente invece la mutazione. In questo caso si ha la fusione delle due blastocisti, quindi l’insieme dei foglietti che deriveranno dalla fusione di queste due blastocisti sarà costituito dal 50% di foglietti normali, derivanti dalla bastocisti normale, e dal 50%di foglietti anomali perché derivanti dalla blastocusti mutata. I genomi saranno per il 50% normali e per il 50% alterati; l’individuo a sua volta sarà un miscuglio di foglietti normali e non. Questa situazione ovviamente è più grave di quella del mosaicismo. Nella chimera i due zigoti, che dovrebbero dare origine a due individui, si fondono. La gravità dipende dal tipo di mutazione, non necessariamente porta alla morte. Può determinare semplicemente una maggior suscettibilità ad un farmaco, per esempio. Il secondo tipo di mutazioni cromosomiche riguarda la struttura del cromosoma e in questo caso si possono avere delle rotture del cromosoma che possono riguardare un solo cromosoma o più cromosomi. In questo caso le mutazioni vengono suddivise in quattro categorie. La prima categoria riguarda una rottura su un solo cromosoma; la seconda due rotture su un solo cromosoma e possono essere su uno stesso braccio o su due bracci diversi; nella terza categoria si hanno due rotture su due cromosomi diversi; la quarta è caratterizzata da tre rotture di cui almeno due sullo stesso cromosoma. Questo è l’esempio di quello che avviene quando si verifica una sola rottura su uno stesso cromosoma. In questo caso abbiamo la rottura in un braccio lungo di un cromosoma e l’effetto di questa rottura sarà la perdita di questo frammento per delezione e quindi si forma un cromosoma più corto. Quindi una rottura su uno stesso cromosoma porta alla delezione del frammento. Se invece le rotture si trovano una su un cromosoma, una su un altro, si ha l’evento della traslocazione e quindi uno scambio di materiale genetico tra i due cromosomi, quindi i due frammenti dei due cromosomi vengono deleti entrambi. Poiché però il genoma tende alla stabilità, si ha la fusione di questi frammenti con il cromosoma più piccolo che è rimasto, quindi si hanno dei cromosomi che contengono la parte finale di un cromosoma, e questo invece conterrà la parte finale in un altro cromosoma e questo evento viene detto traslocazione reciproca. In questo caso i due cromosomi derivanti sono bilanciati ed entrambi contengono tutto il corredo genetico precedentemente presente sul primo e sul secondo cromosoma. Nel caso invece in cui le rotture avvengono molto vicino al centromero, quello che si ha è una fusione centrica con la formazione di un cromosoma più lungo, allungato e uno più corto (costituito dal braccio corto del primo cromosoma e dalla parte del braccio corto del secondo cromosoma) che invece viene perso. Quindi la traslocazione è semplicemente uno scambio di materiale genetico che andrebbe altrimenti perso se la www.slidetube.it Pagina 4 rottura avvenisse su un solo cromosoma e questa fusione è dovuta al fatto che i cromosomi tendono alla stabilità. Nel caso in cui invece avvengano due rotture sullo stesso cromosoma, dobbiamo distinguere se le rotture avvengono sullo stesso braccio del cromosoma o sue bracci differenti. In questo caso gi effetti saranno gli stessi con inversione e delezione interstiziale con la formazione del cromosoma ad anello ma con effetti fenotipici diversi. Nel caso in cui le due rotture si trovino sullo stesso braccio si può avere un’inversione del frammento compreso tra le due rotture, inversione significa proprio una rotazione di 180° del frammento compreso tra le due rotture. Quindi mentre nel cromosoma normale abbiamo la regione A che viene prima della regione B partendo (o passando???) dal centromero e andando verso il braccio lungo, in seguito a inversione avremo che la regione B viene prima della A. Questa inversione viene detta inversione paracentrica, poichè non viene coinvolto il centromero. Un altro effetto che si può avere è la delezione del frammento compreso tra le due rotture. Quindi in questo caso si avrà la formazione di un cromosoma più corto e di un frammento acentrico, mancante del centromero che quindi non è trasmissibile. Il frammento acentrico che si è formato può anche formare il cosiddetto cromosoma ad anello chiudendosi su se stesso sempre perché si tende alla stabilità, ma anche questo tipo di cromosoma, mancane del centromero, non può essere trasmesso. Nel caso in cui invece le due rotture si trovino su bracci diversi del cromosoma, anche qui si può avere un’inversione, quindi una rotazione di 180° rispetto al centromero, inversione che viene detta in questo caso pericentrica perchè si ha uno spostamento di quello che è il braccio lungo e corto del cromosoma. Anche qui un altro effetto può essere la delezione interstiziale e la parte tra le due rotture può essere deleta, quindi in questo caso si avrà la formazione di due frammenti acentrici e di un frammento centrico che manca dei telomeri, quindi anche questo per la mancanza di telomeri non può essere trasmesso stabilmente. Un altro evento che può avvenire è che questo frammento centrico si possa chiudere su se stesso e dare origine al cromosoma ad nello. Un altro evento che può avvenire è che questo frammento centrico si possa richiudere su se stesso dando origine al cromosoma ad anello. Allora questo è un altro lucido in cui ci fa vedere sempre nell'esempio A gli effetti di due rotture su due cromosomi diversi con la formazione di un cromosoma di fusione dicentrico in questo caso poiché la rottura si ha nel braccio corto quindi si avrà la formazione di un frammento acentrico, mancante di centromero, e un cromosoma invece che presenta due centromeri. Nel secondo caso, si ha l'esempio di tre rotture: due che avvengono su un cromosoma e l'altra sul braccio corto del terzo cromosoma. In questo caso quello che può avvenire è una delezione interstiziale nel primo cromosoma, quindi la regione compresa tra le due rotture A e B viene deleta e si va ad inserire in corrispondenza del sito di rottura C sul secondo cromosoma, il quale subisce così una traslocazione interstizionale, quindi quello che si ha è la formazione di due cromosomi e non vi è qui perdita di materiale genetico. Se le tre rotture invece si trovano all'interno dello stesso cromosoma, una sul braccio corto e l'altro sul braccio lungo quello che si può avere è che il frammento deleto A- B si va ad inserire in corrispondenza della frattura C che avviene sul braccio corto del cromosoma e in questo caso si parla di traslocazione interstiziale e anche qui ovviamente non c'è perdita del materiale genico. DOMANDE mi dispiace ma incomprensibili come le risposte!!!:-( www.slidetube.it Pagina 5 Le sequenze geniche sul cromosoma sono posizionate in modo tale che possono influenzarsi anche tra loro, quindi comunque ci sono delle alterazioni di quella che è la funzione originale della proteina che viene prodotta. Questo è un altro lucido che porta l'esempio di cosa avviene se ci sono due rotture sullo stesso cromosoma è il caso di due rotture su un solo braccio con la formazione di una delezione interstiziale che porta ad un cromosoma più corto oppure di un inversione paracentrica questa due rotture su bracci differenti che porta alla formazione di un cromosoma più lungo in seguito ad inversione pericentrica e ad un cromosoma ad anello. Insomma questo è un po' un riassunto di quello che abbiamo visto precedentemente. Allora questo è il caso di un individuo che presenta un assetto cromosomico anomalo ma bilanciato in quanto è avvenuta un' aspirzione che è una traslocazione di materiale genetico da un cromosoma all'altro, quindi anche se l'individuo ha un asseto bilanciato presenta fenotipicamente delle caratteristiche che possono essere individuabili da un genetista esperto ma che dalle altre persone comuni non possono essere identificate appunto perché è un individuo normale, anche se genotipicamente presenta questa aspirazione. I gameti che si formano alla meiosi possono essere: si formerà una coppia di gameti bilanciata, i primi due, mentre una coppia di gameti è sbilanciata questo perché il cromosoma più piccolo presenterà parte di materiale genetico che apparteneva all'altro cromosoma quindi in questo caso in seguito alla fertilizzazione di un gamete normale lo zigote che ne risulta è uno zigote con una parziale trisomia dovuta alla presenza di materiale genetico dell'altro cromosoma e una parziale monosomia dovuta ad un unico cromosoma piccolo normale. Questo qui non è un cromosoma normale perché questa parte qui prima della traslocazione faceva parte dell'altro cromosama, quindi alla fine questo cromosoma è come se fosse presente in triplice copia, mentre questo è come se fosse presente in una sola copia, cioè c'è soltanto questo cromosoma piccolo più metà quindi è per questo che si parla di parziale monosomia e parziale trisomia. La stessa cosa avviene in seguito alla fertilizzazione di un gamete normale con quest'altro gamete, anche qui si avrà una parziale monosomia, in questo caso del cromosoam più lungo e una parziale trisomia del cromosoma più corto proprio perché questo pezzettino, questo materiale genetico che è presente sul cromosoma più lungo deriva dal cromosoma più piccolo nella traslocazione che era avvenuta a monte. Mentre i due gameti bilanciati che vengono prodotti in seguito a fertilizzazione daranno origine a zigoti normali, il primo zigote è anche genotipicamente normale perché presenta la doppia copia di ciascun cromosoma, l'altro invece è normale ma ... in quanto la presenza di entrambe le coppie di cromosomi. Questo invece è l'esempio di un soggetto che ha la presenza di un cromosoma di fusione dicentrico identificato come 14-21 quindi c'è un cromosoma 14, un cromosoma 21 normale e il cromosoma di fusione 14-21. I gameti che vengono sviluppati secondo la meiosi possono essere di questo tipo quindi ci sono dei gameti normali nel primo caso in cui sono presenti il cromosoma 14 e il cromosoma 21, un gamete bilanciato perché è presente il cromosoma di fusione e anche qui un gamete bilanciato e un gamete invece che ha solo il cromosoma 21 e anche qui è anormale e si sviluppa poi in un fenotipo patogenico e anche questi sono invece un gamete normale e una gamete anomalo. In seguito a fertilizzazione con un gamete normale quello che si ha è la formazione di uno zigote normale con una coppia di cromosomi 14 e una coppia di cromosomi 21, uno zigote bilanciato con la presenza di un cromosoma 21, un cromosoma 14 e il cromosoma di fusione. Un soggetto affetto da trisomia 14, un soggetto affetto da monosomia 14 e dei soggetti affetti da www.slidetube.it Pagina 6 monosomia o trisomia 21 quindi comunque in questo caso si ha una percentuale di figli di progenie mutata superiore a quello che si è visto precedentemente in cui si partiva da un soggetto che presentava un genotipo equilibrato. Questo è un esempio di un cariotipo femminile in cui i differenti cromosomi sono distinguibili l'uno dall'altro per i differenti bandeggi, che avvengono in seguito all'utilizzo di coloranti come ad esempio il Gensan che va a colorare delle regioni specifiche all'interno del cromosoma e appunto una colorazione che è specifica che permette il riconoscimento e la distinzione dei vari cromosomi l'uno dall'altro. Questo invece è il cariotipo di un soggetto affetto da trisomia 21, quindi un soggetto maschio poiché c'è la presenza del cromosoma Y ed è evidente la presenza di tre cromosomi 21. Di solito quando si chiede un referto di cariotipo oltre al cariotipo viene indicata un'eventuale mutazione che magari può essere presente e il metodo convenzionale per indicarlo è questo: viene prima indicato il numero di cromosomi, viene poi indicata la coppia di cromosomi sessuali quindi XY o XX e in seguito viene poi descritta l'anomalia che è eventualmente presente nel soggetto. In questo caso l'anomalia è una transizione indicata con T e coinvolge i due cromosomi che vengono riportati in parentesi e cioè il cromosoma 9 e il cromosoma 22, quindi c'è stata una traslocazione di materiale genetico tra i due cromosomi, nella parentesi succesiva vengono identificate e descritte le bande coinvolte, quindi in questo caso le bande coinvolte risiedono entrambe nel braccio corto, identificato con Q, di entrambi i cromosomi e riguardano i geni 34 e 11. Per quanto riguarda la simbologia che viene utilizzata per descrivere le singole anomalie avete questa tabella in cui appunto la traslocazione viene indicata con la T, viene anche indicata la natura in cui noi la rivediamo, la delezione con DE l'inversione con INV, la presenza di un sito fragile, suscettibile quindi con la rottura, viene indicata con FRA, il sito eterocromico con H e la presenza di un'eventuale cromosoma ad anello viene indicata con R. Questo cariotipo che vi ho fatto vedere presenta la tipica anomalia che si ha nella leucemia mieloide cronica. Velocemente, è importante proprio per la presenza di queste mutazioni a livello sia della sequenza di DNA che dei cromosomi che a livello dei vari tratti del genoma il controllo dell'espressione genica che si attua in questi quattro punti cioè a livello trascrizionale, post-trascrizionale, traduzionale e post-traduzionale. Questo è un riassunto di quelle che sono le modificazioni e i controlli a questi quattro livelli quindi potete utilizzarla per riguardare quelle nozioni che avete già acquisito. www.slidetube.it Pagina 7 Ora passiamo a quelle che sono le patologie e le malattie che derivano da mutazioni che coinvolgono i singoli geni o i cromosomi. Alle estremità di questo triangolo sono situate quelle che si pensava un tempo potessero essere le cause fondamentali e tutt'oggi possono essere le cause fondamentali delle malattie. Abbiamo delle malattie che possono essere causate da un singolo gene, le cosiddette malattie mendeliane, le quali si è scoperto ormai da una decina di anni che non sono solo dipendenti al 100% dal gene ma sono anche influenzate da quello che è l'ambiente. L'ambiente è inteso, come abbiamo visto nella seconda diapositiva, come fattori ambientali quindi le radiazioni, gli agenti chimici, ecc. Questo perché anche se un individuo presenta una mutazione in un particolare gene non è detto che questa malattia in una determinata situazione, in un determinato ambiente, sotto l'effetto di stimoli ambientali possa manifestarsi. Ad esempio, un soggetto può essere predisposto ad ipertensione ma se non sottoposto a stress continui o comunque se conduce una vita sana non è detto che possa manifestare la patologia, quindi l'effetto dell'ambiente incide sempre su quello che è anche una malattia genetica ed è per questo che appunto non si può dire che una malattia è al 100% genetica così come non si può dire che una malattia sia al 100% causata soltanto da fattori ambientali. In generale possiamo distinguere le malattie su base genica in: difetti a trasmissione mendeliana, che dipendono da un singolo gene; difetti da singolo gene, ma non a trasmissione mendeliana, quindi non classica (espansioni di triplette, mutazioni in geni mitocondriali, difetti del DNA genomico, i difetti multifattoriali e i difetti complessi o citogenetici o cromosomici). Per quanto riguarda le prime due categorie queste le trovate sul libro e dei difetti multifattoriali ne parlerà poi in modo più approfondito il professore. Difetti a trasmissione non mendeliana I difetti a trasmissione non mendeliana possono essere dovuti all'espansione di triplette, per esempio la sindrome dell' X fragile, quindi triplette che possono essere presenti in un numero di copie molto superiore rispetto all'allele normale quindi invece che essere in numero di 50 possono essere comprese tra 200 e 500 copie, mutazioni in geni mitocondriali, da ricordare che i geni mitocondriali vengono ereditati soltanto dalla madre, e in geni di imprinting genomico. Malattie genetiche a trasmissione mendeliana Questo lucido è un po' superato in quanto ad oggi si conosce un numero enormemente superiore rispetto a 4500 geni coinvolti nelle malattie, oggi si aggira intorno ai 7000-8000, questo perché ogni individuo è portatore di più di 5 geni alterati, di solito un individuo ha un numero di geni alterati compreso tra 5 e 8, sono ovviamente delle mutazioni recessive e quindi fenotipicamente non si manifestano se non nel caso di un genotipo omozigote. L'80-85% di queste mutazioni sono familiari, le restanti invece sono delle nuove mutazioni che possono avvenire durante la formazione dello zigote oppure anche durante la vita dell'individuo stesso. I geni possono essere recessivi o www.slidetube.it Pagina 8 dominanti e quindi determinano un'espressività diversa a seconda se sono in eterozigoti o in omozigoti. Per quanto riguarda i geni recessivi si ha una parziale manifestazione in eterozigosi e una completa manifestazione in omozigosi, è possibile avere anche due alleli codominanti, è il caso delle molecole HLA dei gruppi sanguigni, si ha la presenza del preiotrismo di estrizione (???) ovvero un gene che può dare origine a diversi effetti e l'eterogeneità invece genetiche in cui una mutazione di diversi loci dà origine allo stesso effetto. Modelli di trasmissione mendeliana Questi si dividono in: autosomico dominante; autosomico recessivo. Nel caso di un modello autosomico dominante avremo che i soggetti affetti saranno sia quelli che presentano l'eterozigosi sia quelli che presentano l'omozogosi, in quanto c'è l'allele dominante che anche in eterozigosi fa sì che la patologia venga espressa. In questo tipo di trasmissione sono frequenti le insorgenze di nuove mutazioni e frequente è soprattutto l'insorgenza ritardata ovvero nella prole molto spesso la patologia si manifesta in un'età più tardiva rispetto a quella di insorgenza dei genitori. I geni che sono prevalentemente coinvolti in questo tipo di trasmissione sono delle proteine non enzimatiche quindi che sono coinvolte nella regolazione di vie metaboliche quali ad esempio i recettori di membrana o proteine di trasporto oppure delle proteine strutturali. In questa tabella vengono riportati quelli che sono i difetti dei corrispondenti sistemi delle malattie mendeliane autosomico dominanti, ad esempio l'ipercolesterolemia familiare che coinvolge il sistema metabolico, la sindrome di Marsan del sistema scheletrico e così via. Questa tabella la potete leggere e soprattutto approfondire poi sul libro quelle che sono le varie patologie. Questo è un quadro di Pannet per quanto riguarda l'eredità autosomica dominante. Nel caso in cui si ha un genitore che è eterozigote per l'allele dominante e un genitore invece che è normale il 50% della progenie presenterà la malattia e il 50% sarà normale, malattia che si manifesta appunto anche in eterozigoti. Passiamo ora alle malattie mendeliane autosomiche recessive, in questo caso non si manifesta nei genitori se sono eterozigoti mentre coinvolge il 25% della progenie nel caso in cui i due genitori eterozigoti si incontrino tra loro. C'è una bassa frequenza nella popolazione, queste malattia ci sono soprattutto nei matrimoni tra consanguinei proprio perché le mutazioni recessive presenti nei soggetti sono per l'85% ereditarie quindi è più facile che si trovino tra soggetti consanguinei. Presentano un'espressione uniforme, un'insorgenza precoce e sono rare le nuove mutazioni cliniche. Queste mutazioni si ritrovano in geni che codificano per proteine enzimatiche e quasi tutti portano a degli errori congeniti del metabolismo. Qui c'è una tabella dove vengono elencati quelli che sono i www.slidetube.it Pagina 9 difetti con i relativi sistemi, per esempio, la fibrosi cistica per il sistema metabolico, la fenilchetonuria, la galattosemia, ecc. Questi sono altri quadri di Pannet caratteristici dell'ereditarietà autosomica recessiva in cui ci sono tutti i possibili incroci, quindi, incroci fra genitori entrambi eterozigoti, fra un genitore eterozigote e uno sano, tra un omozigote e uno sano e tra un omozigote e un eterozigote. Questo è importante che ve lo rivediate perché viene anche chiesto all'esame. Per quanto riguarda invece le malattie legate alle mutazioni che avvengono nei cromosomi, quindi alle mutazioni cromosomiche abbiamo le malattie a trasmissione legate al cromosoma X. Tutte le malattie legate al sesso sono dipendenti dal cromosoma X perché il cromosoma Y contiene dei geni che codificano per proteine coinvolte nella spermatogenesi quindi la patologia che si può avere è una sterilità e chiaramente può essere tramandata soltanto ai maschi quindi per il momento non ce ne occuperemo. In questo caso i geni possono essere a trasmissione recessiva o dominante, di solito sono recessive anche perché il cromosoma X non si appaia con il cromosoma Y se non per pochissime regioni. Anche qui c'è una tabella con un elenco dei difetti che coinvolgono il cromosoma X quindi la sindrome dell' X fragile e la distrofia muscolare di Duchenne. Questi sono dei quadri di Punnet relativi all'ereditarietà legata al cromosoma X, recesiva. Questa è importante che la rivediate e sappiate poi esporla. Una delle malattie legate al cromosoma X è la sindrome di Turner che è dovuta ad una monosomia del cromosoma X poiché, come ho citato precedentemente, il soggetto fenotipicamente femmina manca di un cromosoma X quindi mostra un ipogonadismo con la perdita dei caratteri sessuali secondari con sviluppo di possibili cardiopatie e c'è un ingrossamento nella parte posteriore del collo. Anche questa la trovate sul libro. In questo lucido vengono invece identificati tutti i geni situati sul cromosoma X e quindi i geni la cui mutazione può portare appunto allo sviluppo di malattie legate all'X, quindi anche questo è importante che lo sappiate. Altri tre esempi di malattie cromosomiche legate alla trisomia 21: la sindrome di Down, la sindrome di Edwards e la sindrome di Patau. Anche qui gli effetti sono dovuti principalmente ad una trisomia 21 ma anche ad eventi di traslocazione o a eventi di mosaicismo, anche questi li trovate sul libro, sia quelli che sono i cariotipi associati e sia le manifestazioni cliniche e la patologia. www.slidetube.it Pagina 10 Questo lucido invece riporta quella che è una scala d'importanza relativa alla correlazione dei geni e dei fattori ambientali. Questo lucido risale a circa 10-15 anni fa, oggi le cose sono un po' cambiate, in quanto si ritiene che la maggior parte delle malattie non sia dovuta o esclusivamente per difetti del singolo gene o esclusivamente per fattori ambientali, ovviamente ci sono delle malattie genetiche in cui la mutazione di un gene è influente però fare delle classificazioni così nette non è possibile. Anche questo è un lucido da tener presente per tutto quello che è il discorso di correlazione tra ambiente e gene. Chiaramente tutto dipende da quale sia il gene mutato, però è da tener presente che le malattie mendeliane sono quelle che hanno una percentuale dovuta all'ereditarietà e a un difetto genico molto superiore rispetto all'ambiente perché ci sono patologie, quali ad esempio la sindrome di Down, che indipendentemente dall'ambiente sono delle malattie genetiche e indipendentemente dai fattori ambientali comunque il soggetto svilupperà la patologia. Un'altra cosa da tener presente è il fatto che le malattie ereditarie non sono soltanto malattie pediatriche in quanto possono insorgere anche in età avanzata; le malattie ereditarie non sono uguali a malattie pediatriche, anche durante il corso della vita dell'individuo possono insorgere delle malattie che appunto sono ereditarie. In questa tabella vengono riportati quelli che sono i meccanismi patogenetici nelle malattie da singolo gene. Viene indicato il tipo di proteina o funzione interessata, la lesione molecolare che ne determina e la malattia. Nel caso in cui la mutazione coinvolga degli enzimi come la fenilalanina idrossilasi si ha lo sviluppo della malattia fenilchetonuria, nel caso in cui la mutazione coinvolga dei recettori quali ad esempio quelli per le LDL si ha lo sviluppo di un'ipercolesterolemia familiare e così via. Questa tabella è da studiare per sapere il tipo di proteina che può essere interessata dalla mutazione, la lesione molecolare che ne consegue e la patologia. In quest'ultimo lucido quello che si riporta sono dei meccanismi biochimici negli errori congeniti del metabolismo. Viene riportata una via metabolica normale in cui un substrato iniziale A in seguito all'azione di 3 enzimi, indicati con Alfa, Beta e Gamma, danno origine rispettivamente a degli intermedi B e C e ad un prodotto finale D. Il prodotto finale D svolge un'azione a feedback sull'enzima che sintetizza A nell'intermedio B. Una seconda via metabolica è data dalle freccette tratteggiate, ovvero C può essere convertito negli intermedi minori: X, Y e Z. Nel caso in cui avvenga una mutazione sull'enzima che catalizza X A D (???), quindi che interessa l'enzima Gamma, quello che si può avere è: l'assenza del prodotto finale D, in quanto l'enzima magari non viene prodotto quindi la via metabolica si blocca a livello dell'intermedio C, di conseguenza D non viene prodotto. Un altro effetto è l'accumulo degli intermedi B e C in quanto gli enzimi Alfa e Beta continuano a sintetizzare questi intermedi che potrebbero essere tossici e portare all'insorgere di alcune patologie. Un altro effetto potrebbe essere un'aumentata utilizzazione della via metabolica secondaria in quanto il prodotto intermedio C, non potendo più essere convertito nel prodotto finale D, percorre quest'altra via e quindi si ha un'eccessiva produzione degli intermedi X, Y e Z che anche in questo caso potrebbero essere tossici e causare dei danni proprio perché essendo una via secondaria non è detto che questi intermedi siano necessari in grandi quantità alla cellula. L'assenza inoltre del prodotto finale D causa un blocco di quella che è la regolazione a feedback del prodotto D sull'enzima Alfa e quindi l'enzima Alfa, che veniva bloccato dal prodotto D, continua invece a sintetizzare e a convertire il substrato A nel prodotto intermedio B che si accumula enormemente e www.slidetube.it Pagina 11 può dare origine a degli effetti tossici all'interno della cellula. Qui vengono infine elencate quelli che sono i precursori, gli intermedi, i prodotti di vie alternative che possono essere tossici ad alte concentrazioni; c'è il blocco metabolico con una diminuita produzione del prodotto finale e non c'è inibizione del substrato tossico che deve essere convertito da questi enzimi in substrati che invece non alterano l'omeostasi della cellula. Questo lucido riassume bene l'azione delle mutazioni su un enzima coinvolto nel metabolismo. www.slidetube.it Pagina 12
Scarica