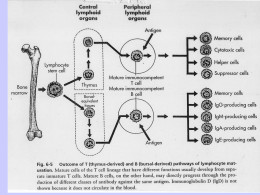
Immunologia Studia tutti quei fenomeni che si verificano nell’organismo in seguito alla penetrazione di un agente estraneo “antigene” e che vede coinvolto il sistema immunitario; un insieme assai numeroso di cellule: i “linfociti” e di molecole: gli ”anticorpi”. Compito del sistema immunitario non è soltanto quello di riconoscere l’agente estraneo giudicandolo non self cioè diverso dalle proprie strutture, ma anche di conservare la memoria dell’avvenuto incontro, così da reagire in maniera più pronta, più efficace e più duratura ad un secondo incontro col medesimo antigene. Le risposte immunitarie Le risposte immunitarie sono di due tipi: umorale e cellulo-mediata Gli antigeni interagiscono con due popolazioni di cellule linfoidi: i linfociti B e i linfociti T. Entrambi derivano dalla cellula staminale del midollo osseo, i primi si differenziano nel midollo osseo, i secondi nel tessuto linfoide del timo. Nei confronti degli antigeni esogeni (batteri o loro prodotti), la risposta immune è di tipo umorale, i protagonisti sono i linfociti B che tramite la cooperazione di specifici linfociti T interagiscono con l’antigene attraverso gli anticorpi molecole ad alta capacità combinatoria che si riversano negli umori circolanti Nei confronti degli antigeni endogeni (antigeni presenti all’interno di una cellula) La risposta immune è di tipo cellulo-mediata, affidata a particolari popolazioni di linfociti T che agiscono sulla cellula infetta, causandone la distruzione. Gli antigeni Si definiscono antigeni tutte quelle sostanze che, introdotte nell’organismo, sono in grado di indurre l’attivazione del sistema immunitario con la produzione di anticorpi o con l’innesco di una reazione cellulo-mediata e di reagire specificatamente con gli anticorpi di cui hanno indotto la produzione o con le cellule effettrici della risposta immunitaria cellulo-mediata Un antigene risulta definito da due principali funzioni: a) l’immunogenicità, cioè la capacità di stimolare il sistema immunitario b) l’antigenicità o specificità di combinazione, ossia la capacità di reagire specificatamente con gli anticorpi o con le cellule di cui ha indotto la formazione. Si definisce aptene una sostanza dotata solo della specificità di combinazione, senza però essere in grado di stimolare il sistema immunitario a meno di non venire introdotta insieme ad un veicolo immunogeno. I requisiti dell’antigene Perché una sostanza possa definirsi antigene, dotata pertanto di immunogenicità e di antigenicità deve possedere i seguenti requisiti: Estraneità, è un concetto fondamentale per l’immunologia che rimanda alla distinzione tra self e non self. Non sempre tuttavia le sostanze estranee sono in grado di stimolare il sistema immunitario e talvolta l’organismo reagisce contro i propri costituenti (malattie autoimmunitarie) Peso molecolare perché una sostanza possa essere immunogena deve avere un peso molecolare opportuno (non inferiore a 1000). Il peso molecolare da solo non basta a definire un antigene, ci sono infatti sostanze che pur avendo un alto peso molecolare non sono antigeni. Complessità molecolare è fondamentale che una sostanza unisca ad un opportuno peso molecolare una certa complessità strutturale, infatti sostanze ad alto p. m. ma con una struttura lineare possono essere debolmente o per nulla immunogeni Stato fisico antigeni in forma aggregata meglio degli antigeni solubili Configurazione sterica antigeni in configurazione L Determinanti antigenici (epitopi) I determinanti antigenici o epitopi sono specifiche regioni poste sulla superficie dell’antigene la cui configurazione sterica può essere riconosciuta dal sistema immunitario. Il numero di epitopi per molecola, cioè la valenza dell’antigene può variare moltissimo. Antigeni come le proteine che possiedono numerosi epitopi diversi sulla superficie appartengono alla classe degli antigeni timo-dipendenti. Essi necessitano di tre tipi di cellule (il linfocito B, il macrofago e i linfociti T helper) per evocare una risposta immunitaria Antigeni per lo più di natura polisaccaridica, che presentano lo stesso epitopo sulla superficie, ripetuto molte volte, sono considerati timo-indipendenti e per indurre la differenziazione del linfocita B a plasmacellula anticorpo produttrice non necessitano del linfocita T helper, ma solamente del linfocita B e del macrofago Fattori che influenzano le risposte immunitarie Dose dell’antigene: le migliori risposte immunitarie si ottengono con dosi ottimali Via di somministrazione: le migliori risposte si ottengono utilizzando vie che portano l’antigene più direttamente a contatto con il sistema immunitario Specie e ceppo dell’animale: la risposta immunitaria varia nell’ambito di specie differenti e in ceppi diversi della stessa specie (controllo genetico risposte immuni) Età e sesso: le migliori risposte si hanno nell’età adulta (massima funzionalità del sistema immunitario), esiste poi differente risposta nei due sessi per l’influenza degli ormoni steroidei Storia immunologica del soggetto: individui che hanno ripetuti contatti con gli antigeni sviluppano risposte più pronte e più efficaci. Anticorpi Sono immunoglobuline specifiche sintetizzate in risposta ad una stimolazione antigenica. Appartengono alla frazione γ globulinica del siero Sottoponendo il siero ad elettroforesi esso si separa in albumina e tre componenti globulinici. Gli anticorpi si trovano nella frazione γ globulinica Poiché, oltre alla frazione γ globulinica, anche altre globuline possono avere funzione anticorpale si è deciso di denominare gli anticorpi: immunoglobuline e di indicarli con la sigla: IgG Struttura degli anticorpi (esperimenti preliminari) Per poter formulare la reale struttura delle immunoglobuline sono state analizzate diverse evidenze sperimentali: 1) Peso molecolare la maggior parte delle molecole ha un peso di ~ 150000 dalton 2) Legami disolfurici nella molecola sono presenti da 15 a 20 legami disolfurici 3) Trattamento con agenti sulfidrilici la rottura selettiva dei legami disolfurici con agenti sulfidrilici determina la comparsa di quantità equimolecolari di due catene peptidiche. Uno dei peptidi pesa 50000 dalton ed è chiamato catena pesante, l’altro 25000 dalton ed è chiamato catena leggera Dal momento che la molecola originale pesa 150000 dalton è chiaramente formata da due catene pesanti e due catene leggere tenute assieme da legami disolfurici Struttura degli anticorpi (digestione enzimatica con papaina) La digestione con l’enzima proteolitico papaina spezza la molecola in due frazioni principali: una frazione (45000 dalton) possiede il sito di legame con l’antigene in sigla Fab, l’altro frammento può essere cristallizzato e viene chiamato frammento cristallizzabile o FC (50000 dalton). Facendo la somma dei pesi molecolari del Fab e di Fc si deduce che la molecola debba contenere due Fab e un Fc Struttura degli anticorpi (trattamento enzimatico con pepsina) Trattando la molecola anticorpale con l’enzima pepsina si ottiene un unico grosso componente (100000 dalton) più dei piccoli peptidi. Il componente più grande è bivalente, presenta un peso molecolare maggiore rispetto a quello dei frammenti Fab ottenuti con papaina ed è chiamato F(ab’)2. Il trattamento con agenti sulfidrilici rompe il legame disolfuro tra le catene e origina due strutture monovalenti indicate come Fab’ Struttura delle immunoglobuline La figura mostra una molecola composta da due catene pesanti (H) di 50000 dalton contenenti 440 aminoacidi ciascuna e due catene leggere (L) di 25000 dalton (220 aminoacidi) legate insieme da legami disolfuro; a seconda del punto in cui viene spezzata la catena pesante risulteranno due frammenti Fab monovalenti e un frammento Fc, oppure un frammento F(ab’)2 e alcuni piccoli peptidi. Si può notare che il frammento F(ab’)2, ottenuto in seguito a digestione con pepsina contiene una quantità maggiore di catene pesanti di quanto non ne contenga il frammento Fab ottenuto attraverso l’idrolisi con papaina. La porzione cerchiata della molecola (estremo aminoterminale) rappresenta il sito di legame con l’antigene. Classi immunoglobuliniche In generale gli anticorpi hanno p.m. 150000 dalton e coefficiente di sedimentazione 7 S. Esaminando preparazioni grezze risulta che gli anticorpi hanno pesi molecolari varianti tra 150000 e 1000000. Le immunoglobuline non sono tutte uguali, ma possono essere divise in 5 classi chiamate: IgG, IgM, IgA, IgD, IgE Le differenze antigeniche tra le classi sono localizzate a livello delle loro catene pesanti. Le catene pesanti sono indicate con le lettere greche corrispondenti: gamma, mu, alfa, delta ed epsilon. Gli studi effettuati sulle catene leggere rivelano che ne esistono 2 tipi principali: kappa e lambda presenti in tutte le cinque classi. La regione cardine o cerniera è la zona di massima flessibilità, è ricca di residui di prolina e assicura pertanto l’apertura e chiusura dei Fab in seguito al legame con l’antigene Classi immunoglobuliniche: IgG Rappresentano più del 70% degli anticorpi umani. Sono formati da 2 catene pesanti (γ) e 2 catene leggere (K o λ). Esaminando la sequenza aminoacidica delle catene leggere si osserva che a cominciare dall’estremo NH2 terminale, metà della sequenza (110 aminoacidi) mostra una variabilità estrema, la seconda metà è invece relativamente costante, queste regioni sono chiamate regione variabile (VL) e regione costante (CL) della catena leggera. Nelle catene pesanti circa 110 aminoacidi della porzione NH2 terminale sono variabili (VH) mentre la rimanente porzione è costante. Poiché la catena pesante è 2 volte più lunga della catena leggera, la regione costante è 3 volte più lunga rispetto alla corrispondente delle catene leggere (CH1, CH2, CH3). Ogni singola regione o dominio contiene un legame SS e un carboidrato ed è associata a specifiche funzioni (capacità di attraversare la placenta, fissare il complemento, legarsi ai fagociti). Le porzioni variabili della catena leggera e della catena pesante formano i siti attivi, cioè le specifiche regione in grado di riconoscere l’antigene. Specificità dell’anticorpo La specificità dell’anticorpo è funzione della struttura spaziale delle regioni variabili delle catene leggere e delle catene pesanti che complessivamente costituiscono il sito di legame con l’antigene Se si confrontano le seguenze aminoacidiche delle regioni variabili di differenti immunoglobuline si trovano zone di alta variabilità. Nelle immuniglobuline umane esistono tre regioni ipervariabili sulla catena leggera e quattro regioni ipervariabili sulla catena pesante E’ stato dimostrato che queste zone di alta variabilità costituiscono il sito di contatto che lega l’anticorpo con il suo antigene, esse sono chiamate: regioni che determinano la complementarietà (CDR) Sottoclassi delle IgG In base alle differenze antigeniche insite nella catena pesante le IgG possono essere suddivise in 4 sottoclassi: IgG1, IgG2, IgG3, IgG4. la ragione per cui queste molecole sono designate come sottoclassi e non come ulteriori classi è dato dal grado di omologia esistente tra i vari tipi antigenici di Ig. Analizzando infatti, le sequenze aminoacidiche delle porzioni costanti delle catene pesanti delle varie sottoclassi si nota che esiste una omologia di più del 90%; mentre invece confrontando le catene pesanti appartenenti a classi differenti, l’omologia non supera il 30%. Le IgG1 rappresentano il 59% delle immunoglobuline totali, le IgG2 il 30%, le IgG3 l’8% e infine le IgG4 il 3%. le differenze tra le sottoclassi vanno ricercate in una differente lunghezza della regione cardine che è sensibilmente più lunga per le IgG3 e per le IgG4 il che le rende più suscettibili alle proteasi e di conseguenza più rare. Classi immunoglobuliniche (IgM) Hanno un peso molecolare di circa 900000 dalton, un coefficiente di sedimentazione di 19 S e sono 5 volte più grandi di una IgG essendo dei pentameri. Ciascuna unità è costituita da due catene pesanti di tipo µ e due catene leggere (K o λ). La catena µ è più pesante della catena γ di una IgG in quanto contiene una regione variabile e 4 regioni costanti (CH1, CH2, CH3, CH4). Una catena peptidica chiamata J lega le 5 unità monomeriche a formare la molecola completa. Le IgM sono i primi anticorpi ad apparire durante la risposta primaria, la loro capacità di legare l’antigene è molto alta (possedendo 10 potenziali siti di legame). In forma monomerica le IgM si trovano sulla superficie delle cellule produttrici di anticorpi e dei loro precursori e in tal modo ne regolano la sintesi quando queste cellule incontrano un appropriato antigene. Classi immunoglobuliniche (IgA) Sono presenti come monomero nel siero. Nelle secrezioni mucose (saliva, lacrime, secrezioni nasali) sono presenti come dimero legato da una catena J e stabilizzato da un componente secretorio prodotto dalle cellule epiteliali. Le IgA sono importanti nel conferire resistenza nei confronti delle infezioni delle mucose, neutralizzando gli antigeni di superficie degli agenti infettanti. Comprendono 2 sottoclassi: IgA1 e IgA2 che differiscono per la natura diversa della catena pesante. Le IgA secretorie sono presenti anche nel latte materno e per tale via possono essere trasferite al neonato contribuendo, insieme alle IgG placentari, alla protezione immunitaria “passiva” del neonato. Classi immunoglobuliniche (IgD) Le IgD rappresentano lo 0,2% circa delle immunoglobuline sieriche Hanno un peso molecolare di 180000 dalton, sono composte da due catene leggere e da due catene pesanti note come catene δ che possiedono un p.m. superiore alle catene γ delle IgG. Originariamente si era postulato che tale peso molecolare fosse dovuto alla presenza di un quinto dominio, ma poi si è dimostrato che la lunghezza della catena era causata da una regione cardine più lunga. Questa regione è suscettibile alla rottura da parte delle proteasi, il che spiega la scarsa concentrazione. La maggior parte delle IgD si ritrova a livello della superficie dei linfociti B, dove svolgono una funzione regolatoria nella sintesi di altre classi immunoglobuliniche. Classi immunoglobuliniche (IgE) Le IgE sono presenti nel siero in modesta concentrazione, sono formate da due catene pesanti chiamate ε, 20000 dalton più pesanti delle catene γ delle IgG e di conseguenza presentano un ulteriore dominio. Le IgE sono responsabili delle allergie pertanto possono trovarsi negli individui allergici in elevata concentrazione. L’ulteriore dominio permette il legame alla superficie delle mastcellule, ciò comporta una reazione che porta alla liberazione di sostanze farmacologicamente attive come istamina e serotonina causa di dilatazione capillare, alterazione della permeabilità e costrizione bronchiale Produzione degli anticorpi: risposta I°e II° Per risposta primaria si intende la produzione di anticorpi quando un antigene penetra per la prima volta in un organismo. Per risposta secondaria si intende la produzione di anticorpi quando lo stesso antigene penetra per la seconda volta nell’organismo. Esiste una sostanziale differenza tra la produzione di anticorpi durante il corso della risposta primaria e durante il corso della risposta secondaria. Andamento cinetico risposta I°e II° Nel corso della risposta I°, gli anticorpi compaiono nel sangue dopo alcuni giorni (fase di latenza) per raggiungere l’apice verso la terza settimana e scomparire intorno alla V°. Le Ig prodotte per prime sono le IgM sostituite poi dalle IgG. Durante questa fase l’organismo produce dei linfociti a lunga vita le “cellule della memoria” che conservano il ricordo dell’ incontro con l’antigene e sono gli effettori della risposta II°. La risposta II° è caratterizzata da un periodo di latenza più breve, dall’essere evocata da piccole dosi di antigene , dal raggiungimento di livelli anticorpali più elevati e rapidi e che si mantengono più a lungo nel tempo. Gli anticorpi inoltre appartengono alla classe delle IgG e sono prodotti dal differenziamento delle cellule della memoria. Reazione antigene-anticorpo La reazione antigene-anticorpo è quella reazione nella quale un antigene reagisce con l’anticorpo corrispondente dando luogo ad un complesso denominato immunocomplesso. La combinazione avviene tra gli epitopi (superficie antigene) e i siti combinatori (anticorpo) La combinazione di un antigene con il rispettivo anticorpo può essere considerata una reazione bimolecolare reversibile accompagnata da una lieve variazione di energia libera, ciò prova che le forze che uniscono l’antigene e l’anticorpo sono forze di legame deboli quali: -forze di Van der Waals (attrazione reciproca tra atomi) -forze di Coulomb (gruppi ionici con cariche opposte) -legami H (interazioni tra gruppi polari non ionici) Stabilità dell’immunocomplesso Poiché le forze che tengono unito l’antigene all’anticorpo sono di tipo debole, la stabilità dell’immunocomplesso dipende dalla formazione di numerosi legami, ciò è possibile solo quando esista notevole coincidenza tra i gruppi reattivi dell’antigene e quelli dell’anticorpo. Deve pertanto esistere complementarietà tra la configurazione sterica del sito combinatorio dell’anticorpo e gli epitopi della molecola antigene. La reazione antigene-anticorpo è una reazione altamente specifica per cui ogni anticorpo si lega esclusivamente al determinante antigenico del quale ha indotto la formazione, discriminandolo anche tra molecole molto simili. Velocità della reazione antigene anticorpo La reazione antigene anticorpo è tra le più rapide reazioni bimolecolari conosciute, considerata la scarsa probabilità che ogni collisione tra antigene e anticorpo possa portare alla formazione dell’ immunocomplesso. La velocità della reazione antigene anticorpo è influenzata da: TEMPERATURA aumentando la temperatura aumenta la velocità della reazione a causa della maggiore energia cinetica impressa alle particelle dell’antigene e dell’anticorpo. PH e CONCENTRAZIONE SALINA DEL MEZZO la presenza di ioni nel mezzo neutralizza le cariche elettriche favorendo l’incontro tra antigene e anticorpo. Le reazioni sierologiche: presupposti L’avvenuta formazione dell’immunocomplesso “in vitro” si accompagna a manifestazioni direttamente apprezzabili ad occhio nudo e pertanto può essere dimostrata. Di conseguenza lo studio della formazione dell’immunocomplesso può essere utilizzato nella pratica a scopo diagnostico mediante le cosidette “reazioni sierologiche” Le reazioni sierologiche La reazione sierologica è quindi una reazione la quale, mettendo a contatto un siero contenente o che si presume possa contenere anticorpi, con un determinato antigene, permette di apprezzare l’avvenuta formazione dell’immunocomplesso I reagenti delle reazioni sierologiche sono due: il siero a contenuto anticorpale noto o presunto e l’antigene. Di questi due reagenti uno deve essere sempre noto. L’avvenuta formazione dell’immunocomplesso, visivamente apprezzabile, ci identifica automaticamente il parametro incognito. Reazioni sierologiche: tipi di reazioni Le reazioni sierologiche sono ambivalenti, esse permettono: a) disponendo di un antigene noto, di dimostrare in un siero la presenza di anticorpi verso quel determinato antigene. b) disponendo di un siero contenente un anticorpo noto, di dimostrare in un materiale la presenza dell’antigene corrispondente. Le manifestazioni macroscopicamente apprezzabili della reazione antigene anticorpo sono: la reazione di precipitazione e la reazione di agglutinazione. Nel primo caso la formazione e la successiva riunione di immunocomplessi si palesa con la formazione di un fine precipitato, nel secondo caso si formerà un agglutinato più grossolano che si deposita al fondo del recipiente. La velocità delle reazioni sarà favorita dai già citati parametri quali: temperatura, presenza di elettroliti, opportuno pH. Reazione di precipitazione La reazione di precipitazione è una reazione sierologica in cui si combinano un antigene macromolecolare solubile con l’anticorpo corrispondente e nella quale la formazione dell’immunocomplesso si appalesa con la formazione di un precipitato insolubile visibile. Per comprendere le cause che rendono possibile la formazione di un precipitato insolubile a partire da immunocomplessi solubili è necessario esaminare lo svolgimento della reazione in relazione alla concentrazione relativa dei reagenti. Se in una serie di provette si distribuisce una quantità costante di siero immune, cioè contenente anticorpi verso un determinato antigene e si aggiungono quantità crescenti di quell’antigene si osserverà che nelle prime provette la quantità di precipitato è scarsa, aumenterà nelle provette centrali per poi tornare a diminuire nelle ultime provette. Curva di precipitazione Riportando in grafico i risultati dell’esperimento precedente si otterrà una curva che presenta 3 zone distinte. Una prima (zona di eccesso di anticorpo), in cui la quantità di precipitato è scarsa, una zona in cui si ha il massimo di anticorpo precipitato (zona di equivalenza) e infine, una zona in cui, pur aumentando la quantità di antigene, l’anticorpo precipitato anziché rimanere costante diminuisce progressivamente (zona di eccesso di antigene) Curva di precipitazione (interpretazione) Ricercando nel sopranatante delle varie miscele di reazione la presenza di antigene o anticorpo liberi, si evidenzia come nella prima parte della curva ci sia presenza di anticorpi liberi, nella seconda zona non siano presenti ne’ antigeni ne’ anticorpi liberi e infine nella terza zona siano presenti solo antigeni liberi. Le prime due fasi della curva sono prevedibili ossia è logico che aumentando la quantità di antigene aumentino gli immuno complessi che precipitano fino a raggiungere la fase di equivalenza con assenza nel sopranatante di antigeni o anticorpi liberi. Meno prevedibile è il comportamento del precipitato nelle provette della terza zona, dove essendo l’antigene aggiunto in eccesso rispetto all’anticorpo ci si aspetterebbe una quantità di precipitato uguale a quello della zona di equivalenza. Poiché nel sopranatante della terza zona non si ritrovano anticorpi liberi, essi sono tutti legati all’antigene pertanto la diminuzione di precipitato non può avere che una spiegazione. Curva di precipitazione: spiegazione La diminuzione del precipitato nella terza zona non può avere che una spiegazione: una parte degli immunocomplessi che si formano in presenza di un eccesso di antigene e in quantità proporzionale, sono formati da immunocomplessi solubili In una miscela formata da anticorpi bivalenti e antigeni multivalenti (molti epitopi) se i reagenti sono in concentrazione elevata essi si riuniscono in immunocomplessi organizzati in reticoli o tralicci tridimensionali in cu le molecole di antigene e anticorpo appaiono alternativamente legate fino a raggiungere la dimensione critica per farle precipitare Aumentando la quantità di antigene nella miscela si formeranno piccoli complessi (una molecola di anticorpo legata a due molecole di antigene) che non potranno incrementare la loro dimensione per l’assenza di anticorpo libero, rimanendo quindi in“soluzione” senza formazione di precipitato. Stechiometria della reazione antigene anticorpo Nella reazione di precipitazione, la formazione dell’immunocomplesso da un punto di vista stechiometrico obbedisce alla legge chimica delle proporzioni multiple e variabili piuttosto che a quella delle proporzioni fisse e costanti. La reazione è infatti condizionata dalla concentrazione relativa dei reagenti, fermo restando che le concentrazioni di antigene e anticorpo debbono essere sufficientemente elevate perché gli immunocomplessi si possano combinare e si possa apprezzare il precipitato. Per tali motivi la reazione di precipitazione è poco utilizzata nella diagnosi delle malattie infettive (necessita di elevate quantità di antigene e anticorpo) ed è poco usata anche per la ricerca di anticorpi nel siero dei soggetti affetti da patologie infettive poiché in questi casi è necessario svelare anche piccole quantità di anticorpi quali si osservano all’inizio del processo infettivo. Applicazioni della reazione di precipitazione La reazione di precipitazione viene generalmente utilizzata con tecniche qualitative nella identificazione di antigeni utilizzando sieri preparati in laboratorio ad alto contenuto anticorpale. Le indicazioni principali riguardano: la medicina legale il settore alimentare identificazione degli antigeni di specie in campioni di sangue o sperma) identificazione degli antigeni di specie nelle carni di insaccati Si usa in questi casi la tecnica di precipitazione zonale o Ring test che consiste nello stratificare, in una protetta capillare, l’antigene sul siero senza mescolare i reagenti. La reazione positiva è data dalla comparsa di un intorbidamento circolare nella zona di contatto tra i due liquidi. Reazione di agglutinazione Mescolando un siero immune specifico (contenente anticorpi specifici) con antigeni costituiti da elementi cellulari, quali batteri o globuli rossi, si formeranno immunocomplessi che si riuniscono in ammassi voluminosi e visibili ad occhio nudo che sedimentano al fondo della provetta. La reazione di agglutinazione pertanto può essere definita come una reazione sierologica in cui un siero immune o presunto tale viene saggiato con un antigene corpuscolato e nella quale la formazione dell’immuno complesso si palesa con la formazione di ammassi voluminosi. Meccanismo della reazione di agglutinazione Il meccanismo che è alla base della formazione degli immunocomplessi nella reazione di agglutinazione è identico a quello che porta alla formazione degli immunocomplessi solubili nella reazione di precipitazione. Ciò che diversifica le due reazioni è dato dalla diversa natura dell’antigene (corpuscolato nel primo caso, solubile nel secondo) e il diverso aspetto del sedimento che sarà più grossolano nella reazione di agglutinazione mentre invece nella reazione di precipitazione apparirà più fine. Nella reazione di agglutinazione la natura corpuscolata e le dimensioni più consistenti delle particelle di antigene renderà la formazione e la riunione degli immunocomplessi non solo più facilmente apprezzabili, ma ne renderà possibile la formazione anche in presenza di concentrazioni assolute dei reagenti di gran lunga inferiori rispetto a quelle necessarie per la reazione di precipitazione. Applicazioni della reazione di agglutinazione La reazione di agglutinazione non richiedendo un esatto proporzionamento dei reagenti e per la sua elevata sensibilità a piccole quantità di anticorpi è usata frequentemente per la ricerca, nel siero di soggetti sospetti di infezione, di anticorpi antibatterici reazione di sierodiagnosi. In questo caso la reazione di agglutinazione viene eseguita con tecnica semiquantitativa allo scopo di stabilire la concentrazione o titolo di anticorpi presenti nel siero in esame. Per l’esecuzione si mescolano in una serie di provette, diluizioni scalari del siero in esame, con una quantità fissa delle sospensioni batteriche e si indica come titolo agglutinante del siero la massima diluizione alla quale si verifica ancora l’agglutinazione. La reazione richiede la presenza di elettroliti la cui funzione è quella di neutralizzare la cariche elettriche ed è favo rita dall’agitazione a bassa velocità e dal riscaldamento a 37° Applicazioni della reazione di agglutinazione La reazione di agglutinazione è usata anche per l’identificazione di antigeni batterici batteriodiagnosi che possono essere presenti in un campione. La reazione si esegue su provetta aggiungendo un anticorpo noto all’antigene incognito, l’avvenuta formazione e riunione degli immunocomplessi si paleserà con la formazione di un agglutinato di aspetto granuloso o fioccoso. La reazione di agglutinazione è inoltre usata per la determinazione del gruppo sanguigno. In questo caso la reazione si esegue su vetrino, mescolando una goccia di sangue con siero immune specifico e osservando l’eventuale comparsa di agglutinazione che sarà evidenziabile entro pochi minuti. Complemento Il complemento (C) è un complesso di diverse proteine: C1, C2…..C9 molte delle quali dotate di attività enzimatica, presente nel siero in forma inattiva. Queste proteine possono venir attivate in maniera sequenziale e ordinata con un meccanismo a cascata nel quale i vari componenti si attivano uno di seguito all’altro e con un meccanismo, definito inoltre, ad amplificazione che porta alla formazione di numerosi componenti alcuni dei quali biologicamente attivi Vie di attivazione del complemento L’attivazione del complemento avviene attraverso due vie: Via classica di attivazione è innescata dal un complesso antigene-anticorpo intervengono tutti i componenti del complemento da C1 a C9 Via alternativa di attivazione è filogeneticamente più antica è indipendente dall’immunocomplesso è innescata da sostanze estranee prodotte da batteri o virus compaiono solo alcuni dei componenti della via classica ma sono presenti componenti specifici Finalità biologiche del complemento L’attivazione del complemento, sia secondo la via classica che secondo la via alternativa porta alla formazione di un complesso litico capace di provocare un danneggiamento della membrana cellulare che causa la lisi della cellula. Il complemento pertanto completa il compito dell’anticorpo il cui ruolo è quello di riconoscere l’agente estraneo e di legarsi ad esso; tuttavia è tale interazione, quella con il complemento, che: Determina la lisi delle cellule estranee (per danneggiamento della membrana cellulare) Facilita la fagocitosi dei microrganismi (attraverso la liberazione di frammenti biologicamente attivi) Causa un’infiammazione locale (stimolando l’attività chemiotattica dei leucociti) Attivazione del complemento (via classica) L’attivazione del complemento secondo la via classica consiste in una serie di reazioni ciascuna delle quali attiva la successiva. L’evento che innesca tale cascata è rappresentato dalla presenza di un immunocomplesso nel quale l’anticorpo deve essere o una IgM o una IgG. L’attivazione del complemento, una volta iniziata, porta alla formazione di un complesso litico che altera irreversibilmente le membrane. L’attivazione del sistema del complemento potrà avere un effetto diverso a seconda della cellula coinvolta nell’immunocomplesso: i batteri Gram- saranno distrutti (presenza della membrana esterna) i batteri Gram+ non possedendo membrana esterna non verranno direttamente lisati ma la liberazione di frammenti attivi a seguito dell’attivazione del complemento ne faciliterà la fagocitosi e di conseguenza l’eliminazione Via classica: unità di riconoscimento Il primo componente della via classica che viene attivato è il C1; esso è diviso in C1q, C1r, C1s. IL C1q esso è una proteina formata da 3 subunità in forma di dimeri ognuna delle quali possiede tre catene (a,b,c) di 200 aminoacidi, terminanti con una estremità globulare capace di legare l’anticorpo a livello degli specifici domini della catena pesante. Una molecola di C1q attraverso le sue 18 catene potrebbe teoricamente legare altrettanti immunocomplessi, in realtà può reagire con un aggregato costituito da un massimo di 13 molecole. L’attivazione del C1q attiva i componenti C1r e C1s (presenti come proteasi inattive). In presenza di Ca++ essi si legano al C1q come complesso tetramerico formato da 2 molecole di C1r e 2 molecole di C1s. Via classica: unità di attivazione Il componente C1s attivato, attiva il componente C4 il quale si scinde in C4a (in soluzio ne) e C4b che si lega alla superficie della membrana, contemporaneamente il componente C2 si scinde in C2a e C2b. Il C2b è eliminato mentre C2a si lega al C4b sulla membrana. Questo complesso chiamato C3 convertasi attiva il componente C3 il quale a sua volta si scinde in C3a e C3b. Il C3a è eliminato in soluzione, mentre il C3b si lega alla C3 convertasi formando il composto C4bC2aC3b chiamato C5 convertasi in grado di attivare il componente C5 che sarà scisso in C5a e C5b. C4a C4b Via classica: unità di attacco La scissione di C5 da parte della C5 convertasi è l’ultima reazione enzimatica coinvolta nella via classica di attivazione del complemento. L’assemblaggio degli altri componenti si svolge spontaneamente. Il C5b attiva il C6 formando C5b-6 e ad esso si aggiungono C7 e C8. Al complesso C5b-6-7-8 che presenta già attività litica, si uniscono da una a tre molecole di C9. Due di questi monomeri dimerizzano per formare il complesso finale di attacco del peso di 1,5 milioni di dalton C5a C5 C5 cov. L’evento litico Il complesso C5b-6-7-8-9(1-3) causa la lisi della membrana cellulare. Secondo la teoria del canale di transmembrana il complesso di attacco formerebbe un canale a forma di ciambella attraverso il quale entrerebbe l’acqua. Il debole della teoria è che la lisi cellulare non è immediata come tale teoria farebbe supporre, pertanto è stata proposta la teoria dello scompaginamento del doppio strato secondo la quale l’evento litico risulta dalla disorganizzazione del doppio strato lipidico, risultato del l’effetto simultaneo e cooperativo di più complessi di attacco adiacenti. Via alternativa di attivazione del complemento La via alternativa di attivazione del complemento chiamata anche via della properdina non richiede la presenza di anticorpi specifici per il suo innesco e pertanto determina un meccanismo di resistenza non specifica e più immediata all’infezione. L’attivazione della via alternativa è mediata da polisaccaridi dell’ospite (LPS), o della parete cellulare dei lieviti, da globuli rossi animali e da membrane povere di acido neuraminico. Questa via inoltre non utilizza i componenti C1, C2, C4 che sono i primi reagenti della via classica ma utilizza dei componenti specifici: il fattore B, il fattore D e la properdina (P). Il risultato generale di questa via è lo stesso della via classica: C3 viene frammentato in C3a e C3b, C5 viene scisso in C5a e C5b permettendo la formazione del complesso che attacca la membrana, identico a quello della via classica. Via alternativa: sequenza di reazioni Normalmente nel plasma esistono piccole quantità di C3b per scissione spontanea di C3. Alcune molecole di C3b si legano al fattore B. Questo in presenza del fattore D si scinde in Ba e Bb. Il composto C3bBb stabilizzato dalla properdina continua a scindere altro C3. Quando al C3bBb si attacca altro C3b, la specificità di tale convertasi cambia e diventa capace di agire da C5 convertasi, scindendo il componente C5. A questo punto la sequenza delle reazioni è uguale a quella della via classica. A livello della scissione del C3 le due vie convergono. C9 Attività biologica delle sostanze formate durante l’attivazione del complemento L’attivazione del complemento comporta la scissione di molecole di C3 e C5 e conseguente formazione di molte molecole di C3a, C3b, C5a. Queste sostanze insieme ad altre che si originano dalla cascata complementare sono dotate di attività biologica e contribuiscono ad ampliare la funzioni del complemento. Anafilotossine I componenti C3a e C5a provocano una anafilassi complemento-indotta e determinano un quadro non diverso da quello della classica anafilassi antigene-anticorpo. Sia C3a che C5a si legano alle membrane delle mastcellule e dei basofili causando il rilascio di mediatori farmacologicamente attivi quali l’istamina e la serotonina il cui maggiore effetto è l’incremento della permeabilità capillare e la costrizione della muscolatura liscia. Tali mediatori contribuiscono all’infiammazione locale. Attività biologica di sostanze formate dall’attivazione del complemento Fattori chemiotattici Ogni sostanza in grado di attirare i leucociti verso una zona di infiammazione viene considerata un agente chemiotattico. Il fattore Ba (via alternativa) e il C5a, privato di un frammento terminale che ne annulla l’azione anafilotossica, sono chemiotattici per PMN e macrofagi, anche il C5b67, parziale complesso litico, ha funzione chemiotattica. Fattori opsonizzanti Il componente C3b è un’opsonina, si lega infatti ad aggregati antigeneanticorpo (presenza di recettori per il C3b) mettendoli a contatto con cellule fagocitarie dal momento che anche quest’ultime contengono recettori specifici per il C3b. Fagocitosi La fagocitosi è uno dei fondamentali meccanismi di difesa antimicrobica dell’organismo, rappresenta l’azione con la quale i fagociti trasportano al loro interno entro vacuoli delimitati da membrana, materiale estraneo in particolare batteri. Le cellule impegnate in questo processo di difesa sono chiamate “fagociti professionali”, sono caratterizzate dalla capacità di accorrere nel luogo di infezione, aderire, inglobare, uccidere e digerire i microrganismi. Possiedono inoltre un recettore per le immunoglobuline (Fc delle IgG) e per il complemento (C3b) Classificazione dei fagociti professionali I fagociti si suddividono in: Sono presenti nel sangue e sono capaci di attraversare Fagociti mobili i vasi e di accorrere nel luogo di infezione. Fagociti fissi Sono presenti a livello dei tessuti, non sono mobili perciò agiscono in sede allorchè il torrente linfatico o ematico porta i batteri a contatto con essi. Fagociti mobili Polimorfonucleati • • • • • • • • Attività fagocitaria molto elevata Vengono mobilitati per primi Sono la prima linea di difesa Fagocitano una sola volta Cellule terminali, vita breve Muoiono dopo aver fagocitato Attaccano microrganismi a localizzazione extracellulare Contengono granuli ricchi di idrolasi, lisozima e altri fattori battericidi Monociti • • • • • • • • Attività inferiore rispetto ai PMN Vengono mobilitati dopo i PMN Rappresentano la II°via difesa Possono fagocitare più volte Hanno vita più lunga, non sono cellule terminali Attaccano parassiti intracellulari obbligati o facoltativi Hanno attività fagocitaria inducibile con segnali come citochine Contengono granuli equivalenti ai PMN Fagociti fissi e fagociti facoltativi Fagociti fissi Sono costituiti dai macrofagi tissutali (derivazione monocitica) Sono presenti nel connettivo, fegato, polmone, milza, linfonodi etc. Agiscono in sede, sono gli antigeni che, trascinati dal sangue o dalla milza vengono portati a contatto con i macrofagi tissutali. Fagociti facoltativi Sono dotati di attività fagocitaria modesta e occasionale Mancano di recettori per le Ig e complemento Sono rappresentati da: fibroblasti, cellule endoteliali, reticolari e spleniche. Fasi della fagocitosi La fagocitosi è un evento complesso che si può suddividere in 5 stadi . • • • • Chemiotassi Opsonizzazione e aderenza Inglobamento e ingestione Uccisione e digestione Esocitosi Chemiotassi La chemiotassi è quel fenomeno per cui cellule mobili quali i PMN e i monociti presenti nel sangue, sono capaci di: 1) avvertire uno stimolo chimico proveniente dal luogo di infezione operato da fattori chemiotattici 2) aderire alle pareti del vaso, 3) attraversarlo con un movimento chiamato diapedesi 4) dirigersi con moto direzionato verso il luogo dell’infezione Fattori chemiotattici • • • • • • Le sostanze chemiotattiche sono numerose e possono agire su tutti i fagociti C5a è il più potente fattore chemiotattico derivato dal complemento C567 il parziale complesso litico svolge azione chemiotattica Leukotriene LTB4 è il più potente fattore chemiotattico generato dalla lipoossigenazione dell’ac. arachidonico nell’anafilassi Ag-Ab e complemento mediata Fattori chemiotattici per eosinofili e neutrofili prodotti da mastcellule e basofili nelle reazioni di ipersensibilità o in seguito ad interazione con anafilotossine nelle reazioni anafilattiche complemento-mediate Formil-metionil peptidi prodotti dagli stessi batteri con azione chemiotattica Il riconoscimento da parte del fagocita di un gradiente di concentrazione della sostanza chemiotattica, rappresenta lo stimolo al movimento direzionato attraverso l’interazione con specifici recettori di superficie. Opsonizzazione Opsonizzazione, letteralmente preparo al pasto, è la capacità di fattori sierici chiamati opsonine di depositarsi sulla superficie dei microrganismi rendendoli più suscettibili alla ingestione da parte dei fagociti. La principale opsonina è rappresentata dal C3b originatosi dalla cascata complementare. I batteri possiedono recettori superficiali per il C3b, l’opsonizzazione pertanto aumenta l’idrofobicità dei microrganismi e ne neutralizza le cariche elettriche superficiali favorendo il contatto con il fagocita Sostanze ad azione antifagocitaria: Capsula (maschera i recettori per il C3b) Proteina M di S. pyogenes (impediscono la fissazione del complemento Proteina A di S. aureus (lega il Fc delle IgG) Tossine citolitiche (azione tossica specifica verso leucociti e macrofagi Fattori che favoriscono la opsonizzazione I fattori che favoriscono la opsonizzazione sono: Anticorpo specifico, Forma con il microrganismo un immunocomplesso che attiva il complemento per via classica Complemento Per attivazione diretta, via alternativa, da parte di sostanze presenti sulla superficie batterica Il batterio in entrambi i casi è ricoperto di C3b, componente che di fatto favorisce la opsonizzazione Ingestione uccisione esocitosi 1)Il batterio opsonizzato viene inglobato tramite pseudopodi e trasportato all’interno del fagocita (fagosoma). 2) La cellula viene racchiusa in un vacuolo. 3)Tramite l’intervento di microtubuli e microfilamenti tende a fondersi con i granuli del fagocita (fogolisosoma). 4)L’ ingestione del batterio genera segnali molto potenti che ne determinano l’uccisione. 5) Il rilascio degli enzimi contenuti nei granuli può avvenire sia all’interno che all’esterno dei granuli in quest’ultimo caso si possono provocare danni ai tessuti circostanti e generare uno stimolo infiammatorio o di cronicizzazione dell’infiammazione. Eventi metabolici correlati alla fagocitosi La presenza del batterio all’interno del fagocita innesca una serie di eventi metabolici quali un rapido consumo di ossigeno che viene ridotto ad anione superossido dal NADPH. Il superossido è convertito ad H2O2 dall’enzima superossidodismutasi. H2O2 insieme alla mieloperossidasi (presente nei granuli) ossida ioni alogeno generando alogenuri ossidati ad alto potere tossico. All’interno del fagosoma unitamente all’azione degli enzimi contenuti nei granuli viene pertanto a generarsi un ambiente altamente sfavorevole per la sopravvivenza della grandissima maggioranza dei microrganismi. I vaccini e il concetto di immunità Per immunità si intende la possibilità di un individuo di risultare “immune” cioè esente da una malattia. Esistono diversi tipi di immunità Immunità naturale di specie, di razza, individuale Attiva da pregresse infezioni naturale Passiva passaggio di ab da madre a feto o attraverso il latte Immunità acquisita artificiale Attiva tramite vaccini Passiva somministrazione di sieri Vaccini I vaccini sono sostanze che introdotte nell’organismo sono in grado di conferire una immunità acquisita artificiale attiva. L’immunità che consegue alla somministrazione dei vaccini è artificiale in quanto gli agenti delle malattie sono introdotti artificialmente, ma è attiva dal momento che l’organismo produce attivamente i fattori di difesa. Con la vaccinazione vengono somministrati gli agenti delle malattie opportunamente modificati e resi apatogeni, si crea pertanto una infezione clinicamente silente ma tale da sollecitare opportunamente i fattori di difesa organici L’immunità che consegue alla somministrazione dei vaccini ha durata variabile da vaccini a vaccino e può essere prolungata con le vaccinazioni di richiamo, dosi di vaccino successive al piano base vaccinale, che possono prolungare anche indefinitivamente il periodo di immunità e con stimoli “secondari” derivanti dalla presenza dei microrganismi nell’ambiente. Tipi di vaccini Gli attuali vaccini sono classificabili in 4 categorie: 1) Vaccini a germi inattivati (uccisi). I microrganismi sono trattati cercando di conservare la loro immunogenicità, questi vaccini tuttavia richiedono la somministrazione di alte dosi per ottenere una buona risposta immunitaria, presentano un lungo periodo di latenza prima della comparsa di protezione e necessitano della somministrazione di frequenti dosi di richiamo. 2) Vaccini a germi vivi e attenuati sono costituiti da germi vivi (varianti apatogene) essi provocano una forma attenuata di malattia capace però di conferire una immunità più efficace e di più lunga durata; richiedono dosi inferiori rispetto al vaccino allestito con germi uccisi. Tipi di vaccini 3) Vaccini allestiti con anatossine, le tossine vengono trasformate in prodotti non tossici (anatossine) con formalina allo 0,5% e calore. Il potere tossico viene eliminato senza alterare la struttura antigene. Questi vaccini inducono un forte stato di immunità, mantenuto con opportuni richiami. 4) Vaccini costituiti da componenti strutturali isolate dai microrganismi possono essere costituiti ad es. dai polisaccaridi capsulari; tali vaccini, essendo costituiti da antigeni timo-indipendenti, danno una immunità di breve durata e non prolungabile con la vaccinazioni di richiamo. Impiego dei vaccini L’utilizzo fondamentale dei vaccini riguarda la prevenzione delle malattie infettive, essi pertanto hanno un impiego preminentemente profilattico. Malgrado l’importante contributo dei vaccini, ancora oggi non è possibile vaccinare contro alcune malattie sia per l’incapacità di mettere a punto un efficace vaccino sia per l’impossibilità di realizzare una vaccinazione di massa, tuttavia molti vaccini tradizionali sono stati in questi ultimi anni migliorati. Esempi: Colera: vaccino ucciso sostituito con vaccino orale ottenuto con DNA ricom. Pertosse: vaccino ucciso sostituito con tossina geneticamente inattivata Tifo: vaccino ucciso sostituito con mutanti apatogeni (vaccino orale) Requisiti dei vaccini: efficacia e sicurezza Un vaccino deve essere efficace, cioè capace di indurre una buona risposta immunitaria, una protezione duratura con un minimo di inoculazioni di richiamo Un vaccino deve essere sicuro cioè innocuo capace di non provocare effetti collaterali di rilievo Questo scopo sarà raggiunto quanto più il vaccino conterrà antigeni “giusti” (antigeni specificatamente correlati alla virulenza) e antigeni “buoni”, (antigeni immunogeni) in grado di sollecitare opportunamente il sistema immunitario. Vaccino ideale e prospettive future Il vaccino ideale è quel vaccino capace di provocare una efficace risposta protettiva, senza effetti collaterali, deve essere chimicamente stabile, di basso costo e di facile somministrazione. I recenti progressi hanno messo a disposizione nuove strategie per la produzione dei vaccini: 1) Vaccini a DNA, il DNA contenente i geni dei fattori di virulenza vengono introdotti nell’organismo e fagocitati inducendo la risposta protettiva 2) Clonazione di DNA ricombinante per la sintesi in cellule lievito di antigeni che inducono una risposta immune proteggente 3) Sintesi chimica di proteine e polipeptidi che mimano i principali antigeni dei microrganismi 4) Costruzione mediante ingegneria genetica di microrganismi mutanti non patogeni 5) Produzione di anticorpi monoclonali mimanti i determinanti superficiali delle proteine di superficie dei microrganismi patogeni Vaccini antivirali Sono usati per la prevenzione delle patologie virali. Sono di due tipi: Vaccini allestiti con virus inattivati Devono essere somministrati in elevata quantità per avere una efficace risposta Vaccini allestiti con virus attenuati o mutanti virali Possono essere somministrati in piccola quantità dal momento che sono vivi; producono uno stimolo antigenico maggiore Prospettive future Per i vaccini virali come per i vaccini batterici, la direttrice fondamentale è quella intesa a isolare e purificare la componenti virali responsabili della immunogenicità e di allestire vaccini usando quanto possibile antigeni prodotti per sintesi o ottenuti in forma pura I vaccini antivirali disponibili in Italia sono: Vaccino antipoliomelitico Vaccino antiepatite B Vaccino antimorbillo Vaccino antiparotite Vaccino antirosolia Vaccino antiinfluenzale Vaccino antirabbico Le vaccinazioni in Italia Obbligatorie Poliomelite, tetano, difterite, epatite B Consigliate Morbillo, pertosse, parotite, Haemophilus influenzae tipo B, rosolia (sesso femminile) Previste per alcune categorie Tifo, tubercolosi, rabbia, varicella Utili in talune circostanze Influenza Abolite vaiolo Immunizzazione passiva: sieri immuni L’immunizzazione passiva consiste nella somministrazione di sieri immuni cioè contenenti anticorpi contro un determinato batterio o una data tossina, a scopo terapeutico Tale tecnica, mette immediatamente a disposizione dell’organismo gli anticorpi di cui ha bisogno e serve a contrastare un’infezione già in atto; i sieri possono avere un’applicazione oltre che terapeutica anche profilattica quando si abbia ragione di ritenere che un individuo è esposto ad un rischio imminente di malattia Attualmente la disponibilità di farmaci antibatterici ha ridotto l’impiego della sieroterapia nelle infezioni, pur tuttavia, l’immunizzazione passiva rimane un presidio fondamentale nelle infezioni da germi esotossici, dove la terapia è rivolta ad impedire il legame tossina-cellula cosa che è ottenibile esclusivamente con l’anticorpo specifico Produzione di sieri immuni e durata dell’immunità I sieri immuni vengono allestiti in animali di grossa taglia sottoposti a trattamento immunizzante con gli antigeni nei confronti dei quali si desidera ottenere produzione di anticorpi, quindi salassati quando il livello anticorpale è massimo. I sieri animali, contenendo tracce di proteine eterologhe, possono causare reazioni di ipersensibilità anche assai gravi. Oggi si impiegano preparazioni di γ globuline iperimmuni ottenute da volontari umani che si sono vaccinati e il cui sangue ha un contenuto anticorpale specifico assai alto. Le preparazioni di gamma globuline umane hanno il vantaggio, rispetto ai sieri animali di non dar luogo a fenomeni allergici. In entrambi i casi, l’immunità che consegue è di breve durata (il tempo richiesto dagli anticorpi per venir metabolizzati cioè 20-30 giorni) Impiego dei sieri immuni I sieri più utilizzati sono: • Sieri antiesotossici (difterite, tetano, butulino, gangrena gassosa) • Siero antivipera o altri veleni animali (scorpioni, ragni, serpenti) • Sieri iperimmuni contro malattie esantematiche (morbillo, rosolia, scarlattina) allo scopo di alleviare i sintomi e prevenire le complicanze. • Siero antirabbico per il trattamento dei soggetti esposti all’infezione, unitamente alla somministrazione di vaccino (dato il periodo di incubazione della malattia assai lungo) Sieri immuni di comune impiego patologia tipo siero impiego commento butulino IGE (animali) terapia somm. precoce difterite IGE profilas. o terap. somm. precoce pertosse IGI (umane) profilas. o terap. riduz. complic. tetano IGI profilassi assoc. con vacc. morbillo IGI profilassi somm. precoce parotite IGI profilassi dubbia efficacia rosolia IGI profilassi prevenz. rischi feto IGE=eterologhe IGI=iperimmuni Ipersensibilità Le reazioni di ipersensibilità sono reazioni che esprimono una aumentata suscettibilità dell’organismo nei confronti di determinati antigeni, piuttosto che un potenziamento della risposta immune Il termine ipersensibilità, pertanto, sta ad indicare una risposta secondaria (penetrazione dell’antigene per la seconda volta) che avviene in forma esagerata o inappropriata o che si risolve in un danno tissutale. Reazioni di ipersensibilità Le reazioni di ipersensibilità sono di due tipi: immediato e ritardato Caratteristica Tipo immediato Tipo ritardato Mediatori Anticorpi Cellule Tempo comparsa reazioni Cellule coinvolte Pochi minuti 24-48 ore PMN e plasmacellule linfociti e macrofagi Tipi di ipersensibilità I fenomeni di ipersensibilità sono divisi in 4 tipi: • • • • Reazioni di tipo I o anafilattiche Reazioni di tipo II o citotossiche Reazioni di tipo III o da immunocomplessi Reazioni di tipo IV o di ipersensibilità ritardata Le reazioni di tipo I, II, III, sono di tipo immediato Le reazioni di tipo IV sono di tipo ritardato Ipersensibilità di tipo I - anafilassi Questo tipo di ipersensibilità è una reazione allergica immediata dovuta alle immunoglobuline E che vengono prodotte, in elevata quantità in taluni individui, in seguito alla stimolazione con particolari antigeni denominati allergeni. I soggetti coinvolti sono una modesta percentuale di popolazione che è sensibile a particolari sostanze e ha la capacità di produrre in gran quantità anticorpi appartenenti alla classe delle IgE. Tra gli allergeni più comuni abbiamo: Antigeni completi come: pollini, epiteli di animali, polveri oppure proteine somministrate come: siero di cavallo od ormoni Apteni sostanze a basso peso molecolare che si legano ai tessuti o alle proteine sieriche, stimolando così la produzione di IgE come taluni farmaci Le fasi dell’anafilassi Le reazioni anafilattiche sono reazioni secondarie che possono essere così schematizzate: 1) 2) 3) 4) 5) Esposizione all’allergene Produzione di anticorpi della classe delle IgE Legame delle IgE alle mast cellule Nuova esposizione all’allergene Legame dell’antigene con le IgE legate alla superficie delle mast cellule 6) Rilascio di mediatori chimici attivi da parte delle mast cellule sensibilizzate 7) Azione di questi mediatori su vari organi N.B. il complemento, tramite le anafilotossine può indurre la degranulazione diretta dei mastociti (anafilassi complemento-mediata) Il legame delle IgE alle mast cellule Le mast cellule e i basofili (forma circolante delle mast cellule) possiedono sulla loro superficie recettori ai quali si legano le IgE attraverso la porzione Fc della catena ε. La quantità di IgE legata alla cellula è proporzionale alle IgE seriche che è più elevata nei soggetti allergici. Le mast cellule contengono mediatori chimici dell’ipersensibilità immediata altamente attivi. La reazione allergica viene scatenata dal ponte che si viene a creare tra una molecola di IgE e l’altra per la presenza dell’antigene. Ciò trasmette un segnale all’interno della cellula che avvia due tipi di reazioni che comportano la liberazione di mediatori farmacologici: neoformati e preformati. Mediatori chimici preformati rilasciati dalle mast cellule sensibilizzate Il legame a ponte, operato dall’antigene, tra due molecole di IgE legate alla superficie di una mast cellula, attiva una serina esterasi citoplasmatica che, tramite l’AMPc, degranula la cellula liberando i mediatori neoformati I principali mediatori neoformati sono: istamina, enzimi proteolitici, eparina e fattori chemiotattici L’istamina ha azione rapida e agisce su diversi bersagli attraverso due tipi di recettori: H1 e H2. Gli H1 si trovano nella muscolatura liscia dei bronchi e dei vasi; la loro stimolazione determina broncocostrizione, vasodilatazione e aumento della permeabilità vasale. I recettori H2 si trovano nello stomaco e a livello cardiaco; la loro stimolazione porta ad aumento della secrezione gastrica e ad azione inotropa e cronotropa. Mediatori chimici neoformati rilasciati dalle mast cellule sensibilizzate L’attivazione della serina esterasi, attraverso una fosfolipasi, rende disponibile l’acido arachidonico della membrana che può essere metabolizzato dagli enzimi: lipossigenasi e ciclossigenasi. Prodotti della via della ciclossigenasi: prostaglandine e trombossani. Effetti biologici: contrazione muscolatura bronchiale, vasodilatazione, aggregazione piastrinica Prodotti della via della lipossigenasi: leucotrieni e il fattore chemiotattico LTB4 Effetti biologici: effetto infiammatorio e contrazione muscoli bronchiali. LTD4 e altri fattori chemiotattici: induzione di neutrofili eosinofili e basofili Reazioni anafilattiche Le reazioni anafilattiche sono la conseguenza della liberazione di mediatori farmacologicamente attivi con effetti locali o sistemici Forma meno grave, nota come “allergia atopica” Si verifica nel 10% della popolazione Anafilassi locale Causata da: pollini, acari, polveri domestiche, alimenti Sintomi: rinite, congiuntivite, asma, febbre da fieno, (secondo la via di introduzione dell’allergene) Anafilassi sistemica Le reazioni sistemiche sono la conseguenza della penetrazione profonda dell’allergene nell’organismo. Estremamente grave Può condurre a morte il soggetto in pochi minuti Anafilassi sistemica In passato era spesso provocata dalla somministrazione di siero eterologo Può essere scatenata da: alimenti, prodotti chimici, farmaci Sintomi: grave ipotensione, edema della glottide Ipersensibilità di tipo II- reazioni citotossiche Sono reazioni spesso dirette contro componenti dell’organismo e quindi definibili come reazioni autoimmuni. Questo tipo di ipersensibilità dipende dal fatto che anticorpi, diretti contro la superficie delle cellule bersaglio, interagiscono con molecole del complemento e con una varietà di cellule effettrici causando danni cellulari L’effetto lesivo può essere di tipo diretto o di tipo indiretto L’azione diretta è dovuta all’interazione tra anticorpi (IgG) e antigeni posti sulla superficie cellulare con attivazione del complemento e lisi cellulare L’azione indiretta è dovuta alla opsonizzazione delle cellule bersaglio che in tal modo sono rese suscettibili all’azione di effettori cellulari: cellule K, piastrine, macrofagi, monociti. Ipersensibilità di tipo II: esempi Reazioni trasfusionali Sono reazioni che si verificano quando, nelle trasfusioni, esista una incompatibilità tra il sangue del donatore e il sangue del ricevente. I soggetti possono essere di gruppo A, B, O, AB Se un soggetto di gruppo O che non possiede alcun antigene sulla superficie dei globuli rossi ma possiede nel siero gli anticorpi anti A e anti B, riceve il sangue da un soggetto di gruppo A; le emazie trasfuse reagiranno con gli anticorpi anti A presenti nel siero e si avrà una reazione emolitica I soggetti di gruppo O saranno donatori universali, in quanto non contengono antigeni sulla superficie dei globuli rossi. I soggetti di gruppo AB saranno accettori universali poiché non possiedono anticorpi anti A o anti B nel siero. Ipersensibilità di tipo II: esempi Malattia emolitica del neonato Si verifica in neonati, la cui madre Rh-, sensibilizzata durante una gravidanza precedente, ha prodotto anticorpi anti Rh+ verso i globuli rossi del figlio. Ad una seconda gravidanza gli anticorpi anti Rh+ presenti nella madre e capaci di attraversare la placenta reagiranno contro le emazie del feto Rh+ determinando la lisi delle emazie. Malattie autoimmuni Ipersensibilità di tipo III reazioni da immunocomplessi Questo tipo di ipersensibilità è chiamata da immunocomplessi. Questi, formati dall’unione antigene-anticorpo, sono generalmente eliminati dall’organismo tramite il sistema reticolo endoteliale; se si formano in notevole quantità, possono causare fenomeni di ipersensibilità attraverso una massiccia attivazione del complemento e la liberazione di fattori attivi sulla permeabilità vascolare, di fattori chemiotattici e di fattori che facilitano l’adesività dei fagociti agli immunocomplessi. Si distinguono due tipi di ipersensibilità Eccesso di anticorpo Eccesso di antigene Reazione di Arthus (reazione da immunocomplessi con eccesso di anticorpi) Agli inizi del 1900 Arthus notò che conigli, inculati settimanalmente per via sottocutanea, con siero di cavallo rispondevano a successive dosi dello stesso antigene con una notevole infiammazione locale. Questa reazione, chiamata dal suo scopritore reazione di Arthus, richiede la presenza di grandi quantità di anticorpi che reagiscono con l’antigene inoculato intradermicamente formando un precipitato che penetra nelle pareti dei vasi sanguigni locali Gli immunocomplessi fissano il complemento con formazione di fattori chemiotattici attivi. I PMN attivati fagocitano i complessi immuni causando liberazione di enzimi lisosomiali, determinando necrosi delle cellule adiacenti e conseguente infiammazione Malattia da siero (reazione da immunocomplessi con eccesso di antigene) Si tratta di una reazione di ipersensibilità che si verifica per inoculazione di siero eterologo Dal momento che tali sieri contengono proteine eterologhe, il soggetto sintetizza anticorpi contro di esse Si generano immunocomplessi solubili con eccesso di antigene che restano in circolo e che a livello capillare precipitano attivando il complemento e determinando fenomeni infiammatori La reazione è simile a quella di Arthus, differenziandosi per il fatto che l’una è localizzata mentre quest’ultima è sistemica. Ipersensibilità di IV tipo (ritardata) L’ipersensibilità di tipo IV include le reazioni cellulo-mediate. Sono reazioni secondarie (penetrazione per la II° volta dell’antigene) determinate da particolari antigeni. I mediatori delle reazioni sono cellule: una particolare categoria di linfociti T. Le manifestazioni infiammatorie avvengono dopo 24-48 h dalla penetrazione, per la seconda volta dell’antigene. Ipersensibilità di IV tipo (esempi) L’esempio più classico di ipersensibilità di IV tipo è rappresentata dall’infezione tubercolare Il Mycobacterium tubercolosis possiede degli antigeni di natura proteica che determinano nell’organismo una condizione di sensibilizzazione allergica di tipo ritardato Ipersensibilità ritardata indotta da Mycobacterium tubercolosis -Il M. tubercolosis penetra nell’organismo e viene fagocitato senza essere distrutto. -Gli antigeni proteici mobilitano i linfociti T i quali, sensibilizzati, si moltiplicano generando un clone di cellule T sensibilizzate. -Ad un secondo contatto con l’antigene, le cellule t producono citochine che determinano una infiammazione e formazione di macrofagi attivati
Scaricare