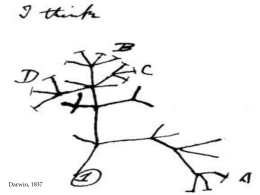
MODELLI EVOLUTIVI Prof. Elisa Prearo LST a.s. 2007/8 Popolazione di organismi molto simili tra loro in grado di accoppiarsi e dare origine a prole fertile. In fase di trasformazione durante il suo movimento nello spazio e nel tempo. Ciò che favorisce il mantenimento di una specie è l’isolamento riproduttivo. SPECIAZIONE: piccoli gruppi isolati riproduttivamente all’interno di una popolazione più ampia possono andare incontro a cambiamenti tali da non permettere più l’interfecondità La specie è : CRITERIO DEFINIZIONE Biologico Un complesso di popolazioni interfeconde, riproduttivamente isolate da tutti gli altri complessi analoghi. NB - Lo scambio genico può essere attuale o potenziale Riproduttivo Un complesso di individui che condividono il medesimo sistema riproduttivo, ed usano segnali visivi, acustici, o chimici per il mutuo riconoscimento. Ecologico Un complesso di popolazioni che occupano una nicchia ecologica propria specifica. Filogenetico Un complesso di popolazioni monofiletiche che evolvono in modo unitario. Tassonomico Un complesso di individui morfologicamente simili fra loro, distinguibili da tutti gli altri complessi analoghi. Nomenclaturale La specie è identificata per mezzo di un binomio latino, di cui il primo termine rappresenta l'affiliazione ad un genere, ed il secondo designa la specie. Modalità di speciazione Si definisce speciazione il processo evolutivo per cui, da una specie originaria derivano due o più specie diverse. La speciazione può essere GRADUALE o IMPROVVISA. Allopatrica Simpatrica Speciazione allopatrica Le popolazioni vivono in territori distanti. L’ostacolo al flusso genico è determinato da separazione geografica GRADUALE MODALITÀ: VICARIANZA - la separazione geografica è dovuta ad eventi geologici (deriva delle placche) che frantumano l’areale della specie (esempi: masse continentali che si allontanano l’una dall’altra; placche che collidono determinando l’insorgere di catene montuose). MIGRAZIONE - la separazione geografica è dovuta ad eventi climatici, che causano la migrazione della specie sul territorio, con conseguente separazione fra popolazioni che migrano in aree diverse. DISPERSIONE - la separazione geografica è dovuta a una migrazione (es.: uccelli) o a trasporto passivo (es.: semi) fortuito a grande distanza (esempio: la colonizzazione delle isole oceaniche). I fringuelli di Darwin: un classico esempio di speciazione allopatrica Speciazione simpatrica Le popolazioni si trovano sullo stesso territorio. L’ ostacolo al flusso genico è determinato da caratteristiche morfologiche, etologiche, o funzionali Una o poche mutazioni possono determinare caratteristiche tali da rendere impossibile od altamente improbabile il flusso genico. La speciazione IMPROVVISA è sempre simpatrica. L’ ostacolo al flusso genico è determinato da una improvvisa mutazione genomica (poliploidismo) Il raddoppio del numero cromosomico determina il blocco, totale od almeno parziale, del flusso genico. Si tratta di un processo del tutto infrequente fra gli animali, ma molto importante nell’evoluzione delle piante Mantenimento dell’isolamento Prezigotico: impedisce l’accoppiamento a. mancanza di opportunità di accoppiamento -isolamento temporale -isolamento ecologico b. incompatibilità di accoppiamento -incomp. sessuale, psicologica o comportamentale -isolamento meccanico -isolamento gametico Postzigotico: impedisce di avere progenie fertile a. non vitalità degli ibridi b. sterilità degli ibridi c. totale disfunzione degli ibridi Modelli evolutivi Evoluzione convergente Si verifica quando due o più gruppi che NON sono strettamente imparentati, evolvono caratteristiche simili. Ciò è generalmente il risultato dell'occupazione di habitat simili e la risposta a pressioni selettive simili. Modelli evolutivi Evoluzione divergente Una specie ancestrale si è evoluta in due specie, che continuano ad evolversi indipendentemente e si differenziano sempre di più nel tempo. Cladogenesi Radiazione adattativa Evoluzione del cavallo: dal genere Eohippus (Hiracotherium) al genere Equus Gradualismo Le teoria neo-darwiniana, secondo la quale l'evoluzione è il risultato dell'accumulo di mutazioni casuali soggette a selezione, implica necessariamente che l'evoluzione sia un processo graduale e più o meno uniforme nel tempo. Questa teoria ha quindi sempre incontrato un convinto sostegno nei genetisti, che possono verificarne la correttezza negli organismi sperimentali (ad esempio il moscerino Drosophila melanogaster, o la pianta annua Erophila verna). La teoria ha sempre destato qualche perplessità nei paleontologi. Infatti, se si osserva la documentazione fossile, non si nota alcuna variazione continua nel tempo. Al contrario: una certa specie compare negli strati fossili di una certa epoca, rimane invariata per alcuni milioni di anni, ed infine scompare, così improvvisamente come era comparsa. Un'altro fatto notevole è che molte specie compaiono contemporaneamente e, dopo un certo tempo (sempre dell'ordine dei milioni di anni) scompaiono contemporaneamente. Dopo un'estinzione di massa, le nuove specie che compaiono non appartengono agli stessi generi ed alle stesse famiglie precedenti l'estinzione, ma rappresentano generi e famiglie nuove. L’'andamento della biodiversità nel tempo: un aumento veloce di diversità, seguito da un periodo di stasi, e poi da un crollo di diversità (estinzione di massa). La storia della vita sulla Terra è tutta costituita da questo andamento, di periodi di "equilibrio" piuttosto lunghi, "punteggiati" da improvvise interruzioni. Equilibri punteggiati Stephen Jay Gould e Niles Eldredge Niles Eldredge e la sua collezione di trombe Su tutti questi dati di osservazione, due paleontologi americani, Stephen J. Gould e Niles Eldredge, hanno proposto una teoria dell'evoluzione che è alternativa rispetto a quella derivata dal darwinismo. Essi asseriscono che: 1. Quando nell'ambiente si manifestano importanti cambiamenti (per esempio, il raffreddamento globale che ha accompagnato le glaciazioni), le specie, non si modificano sostanzialmente: o sopravvivono migrando in altre regioni, o si estinguono. 2. Nei lunghi periodi di equilibrio (quando non vi sono variazioni ambientali drastiche), tutte le nicchie ecologiche sono colonizzate, tutte le specie presenti si diffondono nel massimo grado consentito dall'ambiente, e quindi in ogni specie si manifesta competizione, lotta per la sopravvivenza, e selezione naturale (cioè, i ben noti meccanismi "darwiniani"). Però, in questi periodi non si manifestano grandi novità evolutive: infatti la documentazione fossile ci consegna specie praticamente invariate nel loro fenotipo. 3. Nei brevi periodi in cui avviene un'estinzione di massa, moltissime nicchie ecologiche si liberano. A questo punto, specie che in precedenza occupavano piccole nicchie marginali (e che perciò non hanno lasciato fossili) possono rapidamente occupare le nicchie libere: questo fenomeno, che avviene in presenza di scarsa o nulla competizione, si accompagna con una veloce evoluzione, determinata dal fatto che le condizioni ambientali nuove consentono il manifestarsi e l'evolvere di caratteri diversi rispetto al passato. Nei periodi di veloce radiazione evolutiva, anche organi (o molecole) preesistenti, che svolgevano una certa funzione, possono risultare utili a svolgere funzioni del tutto diverse (si cita, a questo proposito, l'esempio di molecole dalla struttura assai simile fra loro come l'emoglobina del sangue degli animali, i pigmenti della bile, i pigmenti fotosintetici dei Cianobatteri e delle Rodofite, che appaiono derivate da una molecola ancestrale comune, ma svolgono funzioni del tutto diverse in contesti diversi). i neodarwinisti sostengono che la "macroevoluzione" (cioè l'evoluzione di generi, famiglie e phyla diversi) non è altro che lo sviluppo nei tempi lunghi della "microevoluzione" (cioè l'evoluzione che avviene al livello delle popolazioni e delle specie). Ambedue i fenomeni si spiegherebbero allo stesso modo, ricorrendo agli stessi meccanismi. Invece, secondo i sostenitori di questa teoria, la microevoluzione non vale a spiegare la macroevoluzione: si tratterebbe di due fenomeni coesistenti ma diversi. Precisamente, l'evoluzione "darwiniana" (microevoluzione) esiste e si manifesta effettivamente a livello delle popolazioni e delle specie, ma ha sostanzialmente una funzione stabilizzatrice. Invece la macroevoluzione avverrebbe in assenza di competizione e di selezione naturale. Cladogrammi Questo tipo di diagramma riflette le relazioni evolutive degli organismi sulla base della distanza da un antenato comune, con o senza riferimento ad una scala temporale. I cladogrammi sono costruiti sulla base di dati oggettivi, in particolare sulle somiglianze biochimico-molecolari; attualmente si utilizzano le omologie del DNA. La somiglianza delle sequenze di basi del DNA viene utilizzata come misura della distanza evolutiva: tanto più simile è la sequenza del DNA, tanto più vicine evolutivamente sono due specie, poiche' non vi è stato tempo sufficiente perchè si accumulassero molte mutazioni nel DNA a partire dall'antenato comune. Le ramificazioni del cladogramma rappresentano lo sviluppo di una nuova specie. Questo semplice cladogramma mammaliano mostra che i dingo (placentati) e i koala (marupiali) sono evolutivamente più vicini del platipo (monotremi). Questo è un cladogramma più complesso che mostra non solo le relazioni evolutive tra orsi, panda e cani, ma fornisce anche una misura del tempo trascorso dalla divergenza da un antenato comune, condiviso tra le diverse specie. Di sotto è illustrato un cladogramma relativo a sette diversi vertebrati (immagine tratta da Purves et al. , BIOLOGIA, Zanichelli), in cui viene indicato il carattere distintivo emergente nelle divergenze delle diverse classi.
Scaricare