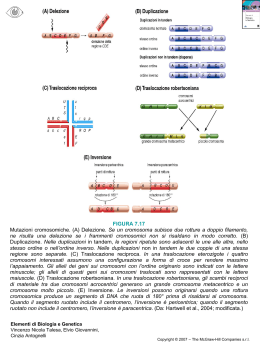
• I meccanismi di controllo del ciclo cellulare normalmente impediscono a cellule con rotture cromosomiche non riparate di entrare nel ciclo cellulare; se il danno non può essere riparato la cellula muore (APOPTOSI). • Gli eventi fisici di rottura e riunione della molecola di DNA che costituisce il cromosoma possono verificarsi spontaneamente oppure essere indotti da un trattamento con radiazioni ad alta energia, come i raggi X e gamma. - Ogni cromosoma è una singola molecola di DNA; - Il primo evento di un riarrangiamento cromosomico è il verificarsi di due o piu’ rottire nel doppio filamento del cromosoma; - Le rotture a doppio filamento sono letali, a meno che non siano riparate; - I sistemi di riparazione congiungono le estremità dei filamenti rimaste libere; - Se i sistemi di riparazione ricongiungono le estremità dello stesso punto di rottura si ripristina la sequenza cromosomica originaria - Se invece vengono unite le estremità di due punti di rottura diversi si verifica un riarrangiamento cromosomico; - I riarrangiamenti cromosomici che sopravvivono alla meiosi sono quelli che danno luogo a molecole di DNA provviste di centromero e due telomeri. Cromosoma acentrico, dicentrico e ponte anafasico - Un altro meccanismo che provoca riarrangiamenti cromosomici è il crossing-over tra segmenti di DNA ripetitivo (duplicato). La presenza di sequenze ripetitive brevi in un cromosoma o su cromosomi diversi causa ambiguità su quali ripetizioni si debbano appaiare tra loro alla meiosi. Se si appaiano sequenze che non sono nella stessa posizione relativa sugli omologhi, il crossin-over puo’ produrre cromosomi aberranti. Delezioni, duplicazioni, inversioni e traslocazioni possono essere tutte causate da tali meccanismi di crossing-over. I riarrangiamenti sono bilanciati e non-bilanciati. A questi ultimi appartengono la delezione (con la perdita di un segmento) e la duplicazione (ripetizione di un segmento di un braccio cromosomico). Le anomalie strutturali insorgono quando le rotture vengono riparate in modo sbagliato. • Un cromosoma senza centromero (acentrico) o con due centromeri (dicentrico) non segrega mai e alla fine verrà perso. • I l d i c e n t r i c o a l l ’ a n a f a s e s a r à t r a s c i n a t o simultaneamente ai poli opposti formando il ponte anafasico e i cromosomi che formano questo ponte non sono incorporati in alcuna cellula della progenie. • I cromosomi con un centromero vengono trasmessi in modo stabile nei successivi cicli anche se strutturalmente anomali • Un altra classe rara di anomalie strutturali è quella degli isocromosomi, si tratta di cromosomi costituiti da due bracci lunghi o corti. Delezione: perdita di un tratto di cromosoma • 1. Conseguenze: Dipendono dai geni o dalle parti di geni che vengono rimosse. 2. Negli individui diploidi gli effetti possono essere attenuati dalla presenza sul cromosoma omologo di una copia dei geni deleti. Identificazione: negli organismi in cui è possibile allestire un cariotipo, le delezioni hanno come conseguenza la formazione di anse non appaiate • Se è deleto l allele dominante in un eterozigote, si ha come conseguenza la manifestazione dell allele recessivo, questa espressione inattesa dell allele recessivo è definita PSEUDODOMINANZA Le delezioni estese (4000kb) possono essere visualizzate mediante le tecniche di bandeggio. Se si esaminano i cromosomi in meiosi di un individuo eterozigote per una delezione, si puo arrivare a determinare la regione deleta in base al fatto che il corrispondente segmento sull omologo normale, non potendo appaiarsi, forma un ansa da delezione. Delezioni terminali Delezioni interstiziali: sono evidenziabili solo con una sonda Una piccola delezione in un gene (delezione intragenica), inattiva il gene coinvolto e si distingue dai cambiamenti di singoli nucleotidi in quanto non puo retromutare. Le delezioni multigeniche eliminano da due a piu geni hanno conseguenze molto piu gravi; se la stessa delezione e presente su entrambi gli omologhi allora la mutazione e quasi sempre letale. Le delezioni producono uno sbilanciamento genico che puo risultare deleterio. La delezione puo essere messa in evidenza mediante analisi citogenetica. Esempi di malattie causate da una delezione Sindrome di Cri-du-Chat (5p-): • Sindrome caratterizzata da un grido acuto del lattante simile ad un gatto, che scompare dopo poche settimane dalla nascita • Basso peso alla nascita e crescita rallentata, microcefalia,volto rotondo • Strabismo, radici nasali allargate • Grave ritardo mentale, difetti cardiaci congeniti La sindrome di Cri-du-chat e provocata da una delezione eterozigote all estremita del braccio corto del cromosoma 5. Tali delezioni si originano per lo piu spontaneamente nelle cellule germinali di un genitore normale del soggetto affetto; quindi, i cromosomi delle cellule somatiche parentali non presentano alcuna delezione. Patologia evidenziabile tramite FISH. La sindrome di Williams è una sindrome autosomica dominante causata da una delezione di un segmento di 1,5 Mb su uno dei due cromosomi 17. L’analisi della sequenza del DNA ha mostrato che il segmento che va incontro a delezione contiene 17 geni non tutti a funzione nota. Il segmento che va incontro a delezione è affiancato da copie ripetute di un gene chiamato PMS, che codifica una proteina coinvolta nella riparazione del DNA. Sequenze ripetute del DNA possono andare incontro a crossing-over ineguali. Questi eventi provocano una duplicazione (che non è osservata) e una delezione che dà origine alla sindrome di Williams. La sindrome di Williams-Beuren è caratterizzata da stenosi aortica sopravalvolare (causata da un deficit di elastina), ritardo mentale associato ad un carattere estremamente socievole ed estroverso anche con gli estranei (quella che è stata descritta come personalità da "cocktail party"), ritardo di crescita spesso associato ad insorgenza postnatale, invecchiamento precoce e un aspetto del volto caratterizzato da tratti grossolani, con palpebre edematose, iride stellata, epicanto dorso nasale depresso e narici antiverse, bocca larga con labbra carnose, guance paffute con mandibola piccola (Facies Elfica). In Italia vi sono circa 3000 casi. Presentano una compromissione all'emisfero destro, difficoltà visivo-spaziali e presentano una dissociazione tra gli aspetti pragmatici, gli aspetti fonologici e sintattici del linguaggio. Presentano un quoziente intellettivo tra 60 e 70. Inversione: • Quando un segmento cromosomico viene exciso e poi reintegrato nel cromosoma dopo rotazione di 180 gradi rispetto all orientamento originale • In generale, non vi è perdita di materiale cromosomico, benchè si possano verificare delle conseguenze fenotipiche quando i punti di rottura sono all interno di un gene o entro regioni che controllano l espressione Inversione: Inversione: • Conseguenze alla meiosi: 1. Se l inversione è in omozigosi la meiosi avviene normalmente senza conseguenze di duplicazioni o delezioni. In eterozigosi i cromosomi omologhi cercano di appaiarsi nel miglior modo possibile; ciò richiede la formazione di anse che comprendono i tratti invertiti. Non vi sono problemi se il corssing-over è assente. Si verificano gravi conseguenze se un crossing-over avviene entro l inversione 2. 3. Inversione pericentrica Inversione pericentrica eterozigote comprende il centromero • L appaiamento degli omologhi richiede la formazione di anse d inversione • Il centromero si trova tra B e C I risultati del crossing-over e delle successive divisioni meiotiche sono due gameti vitali, con i cromosomi non ricombinanti • Gli altri due gameti ricombinanti non sono vitali, perché sbilanciati ( privi di alcune regioni cromosomiche o duplicati) • I soli gameti che possono dare origine a una progenie vitale sono quelli che contengono i cromosomi non coinvolti nell evento di crossing-over • Due crossing-over vicini in un anello d inversione producono quattro gameti vitali Inversione Un individuo è eterozigote per la seguente inversione: A_B_C_D_E_(centromero) F_G_H_I (cromosoma normale); A_B_G_F_(centromero) E_D_C_H_I (cromosoma invertito). Disegnare l’appaiamento dei cromosomi alla meiosi e i quattro cromosomi prodotti se avviene un crossing-over nell’intervallo tra le posizioni G ed F I quattro gameti saranno A_B_C_D_E_(centromero) F_G_H_I (cromosoma normale); A_B_G_F_(centromero) E_D_C_H_I (cromosoma invertito); A_B_C_D_E_(centromero)F_G_A_B (gamete sbilanciato con duplicazione di AB e mancanza di HI); I_H_ G_F_(centromero) E_D_C_H_I (gamete sbilanciato con duplicazione di HI e mancanza di AB). Inversione paracentrica eterozigote • L appaiamento degli omologhi richiede la formazione di anse d inversione • Il crossing-over da origine a un cromosoma dicentrico e ad un frammento acentrico che viene perduto • All anafase il ponte dicentrico si rompe • Dopo la seconda divisione meiotica, due gameti sono vitali, possiedono una serie completa di geni • Gli altri due non sono vitali, perché sbilanciati e privi di alcune regioni cromosomiche • I soli gameti che possono dare origine a una progenie vitale sono quelli che contengono i cromosomi non coinvolti nell evento di crssing-over Inversione • Un individuo è eterozigote per la seguente inversione paracentrica (n0n comprende il centromero): • A (centromero) B_C_D_E_F_G_H_I (cromosoma normale); • A(centromero) B_G_F_E_D_C_H_I (cromosoma invertito). • Disegnare l’appaiamento dei cromosomi alla meiosi e i quattro cromosomi prodotti se avviene un crossing-over nell’intervallo tra le posizioni G ed F Inversione • Una pianta di pomodoro è eterozigote in cis per i tre geni A, B e C (nell’ordine). Le distanze di mappa sono: A-B 12 U.M, B-C 9 U.M. Questa pianta è inoltre eterozigote per un’inversione della lunghezza di 5 U.M che comprende il gene B, ma non comprende A, C o il centromero. Mostrare l’appaiamento meiotico in questa pianta e indicare gli scambi che permettono di recuperare i seguenti gameti ricombinanti: (1) aBC (2) aBc. Qual’è la frequenza attesa dei gameti aC? • • Il gamete aBC deriva da uno scambio tra il gene A e l’inizio dell’inversione. (2) il gamete aBc deriva da due scambi, uno tra A e B e l’altro tra BC, che devono essere o ambedue all’interno o ambedue all’esterno dell’inversione. La frequenza di ricombinazione tra A e C nella pianta eterozigote per l’inversione diventa 12 + 9 –5 = 16 U.M. La frequenza attesa dei gameti aC è quindi l’8%.
Scaricare