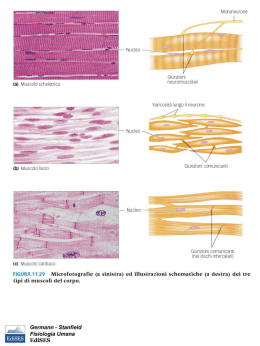
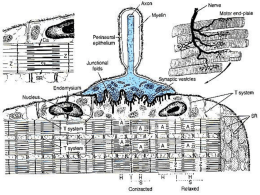
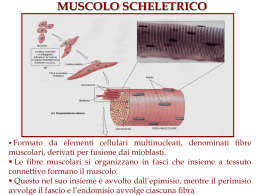
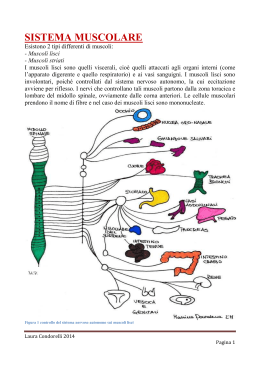
Il muscolo scheletrico • • • • contrattilità: capacità di contrarsi con forza eccitabilità: capacità di rispondere agli stimoli estensibilità: capacità di essere stirato elasticità: capacità di tornare alla lunghezza iniziale. Struttura dell’apparato contrattile I filamenti muscolari • Filamenti spessi: miosina (catene pesanti e catene leggere) sito di legame per l’actina sito si idrolisi dell’ATP (miosina ATPasi) • Filamenti sottili: actina (actina G e actina F) siti di legame per la miosina tropomiosina troponina (troponine T, I e C) L’organizzazione dei filamenti nel sarcomero • • • • • • La struttura delineata da due linee Z costituisce il sarcomero (unità funzionale dei muscoli) Banda A: contiene filamenti spessi di miosina Bande I: sono formate dai filamenti sottili di actina, dalle proteine filamentose associate alle linee Z Linee Z: delimitano le estremità del sarcomero Zona H Linea M: contiene proteine che tengono unite le parti centrali dei filamenti spessi I tubuli trasversi ed il reticolo sarcoplasmatico • I tubuli trasversi: permettono il propagarsi della depolarizzazizione dalla superficie verso l’interno della cellula; stabiliscono contatto con le cisterne terminali del reticolo sarcoplasmatico e contengono una proteina voltaggio-dipendente (recettore diidropiridinico) • Il reticolo sarcoplasmatico: costituisce il punto di immagazzinamento e di rilascio degli ioni Ca2+ per l’accoppiamento eccitazione-contrazione; il reticolo sarcoplasmatico è fornito di un canale per la liberazione degli ioni Ca2+ (recettore per la rianodina) • Gli ioni Ca2+ si accumulano nel reticolo sarcoplasmatico mediante l’azione di una pompa Ca2+/ATPasi • Nel sistema sarcoplasmatico gli ioni Ca2+ si legano alla calsequestrina I recettori diidropiridinico e rianodinico IL CICLO DEI PONTI TRASVERSALI. Una miscela di proteine contrattili in vitro cambia conformazione (si aggrega: actina + miosina = actomiosina) in presenza di Ca+2 e si disaggrega se viene rimosso il Ca+2 e aggiunto ATP. In assenza di ATP il complesso actomiosinico è stabile (complesso del rigor). La pompa Ca2+/ATPasi L’accoppiamento eccitazionecontrazione nel muscolo scheletrico Evoluzione degli eventi nell’accoppiamento eccitazione-contrazione nel muscolo scheletrico • L’accoppiamento eccitazione-contrazione è il meccanismo che media la trasformazione del potenziale d’azione in produzione di tensione • Il potenziale d’azione precede sempre l’aumento della concentrazione intracellulare di ioni calcio che, a sua volta, precede sempre la contrazione Le basi molecolari della contrazione nel muscolo scheletrico 1. 2. 3. 4. 5. I potenziali d’azione della membrana della cellula muscolare si propagano ai tubuli a T. La depolarizzazione dei tubuli a T provoca un cambiamento di conformazione nel recettore diidropiridinico, determinando l’apertura dei canali del reticolo sarcoplasmatico che lasciano passare gli ioni calcio a livello del recettore rianodinico Aumento della concentrazione intracellulare (10-6 M) di calcio Gli ioni calcio si legano alla troponina C con modificazione della conformazione del complesso troponinico Il cambiamento di conformazione del complesso troponinico sposta verso l’esterno la tropomiosina e conseguente interazione tra miosina e actina 6. 7. 8. 9. L’actina e la miosina si legano formando i ponti trasversi. Le teste della miosina impegnate nel ponte trasverso si inclinano facendo scorrere i filamenti sottili e spessi gli uni sugli altri con un movimento a cerniera e producendo così sviluppo di tensione L’evento meccanico che viene definito come scossa produce quindi una tensione che è proporzionale al numero di ponti trasversi formati Il legame actina-miosina attiva la miosina ATPasi conidrolisi dell’ATP. Infine, quando i ponti trasversi si rompono, una nuova molecola di ATP si lega alla testa della miosina e comincia un nuovo ciclo Si ha rilasciamento quando gli ioni calcio vengono riaccumulati nel reticolo sarcoplasmatico (Ca2+/ATPasi). Quando la concentrazione intracellulare degli ioni calcio diviene <10-7 M non vi è più un numero sufficiente di questi ioni per legarsi alla troponina C. Anche in questa fase di rilasciamento l’ATP viene idrolizzato in conseguenza dell’attività della Ca2+/ATPasi Le tappe del meccanismo dell’accoppiamento eccitazionecontrazione Teoria dello scorrimento dei filamenti • Durante la contrazione i filamenti spessi e sottili sovrapposti slittano gli uni sugli altri in un processo energia dipendente, a causa del movimento dei ponti trasversali tra actina e miosina Biofisica del sistema contrattile • I muscoli possono essere rappresentati da due elementi disposti in serie: - un elemento contrattile - un elemento elastico • L’elemento elastico è costituito da una componente passiva (tendine e formazioni connettivali) ed una attiva (ponti trasversali) Relazione lunghezza-tensione • Effetto della lunghezza della fibra muscolare sulla tensione che la fibra stessa può sviluppare • La tensione può essere determinata durante una contrazione isometrica nella quale il muscolo può sviluppare tensione partendo da una lunghezza prestabilita (precarico), ma non può accorciarsi La tensione attiva massimale si sviluppa in corrispondenza della lunghezza muscolare alla quale si verifica una sovrapposizione ottimale dei filamenti sottili e spessi che consente la formazione del maggior numero possibile di ponti trasversali Relazione forza-velocità • • • • • Effetto della velocità di accorciamento in funzione della variazione della forza contro cui il muscolo deve contrarsi, quando cioè il postcarico viene a variare La relazione forza-velocità può essere determinata lasciando che il muscolo si accorci. In questa situazione è la forza ad essere fissata e si ha quindi una contrazione di tipo isotonico La velocità dell’accorciamento riflette la velocità alla quale si formano i ponti trasversali La velocità di accorciamento è massima (Vmax)quando il postcarico è zero La potenza è il prodotto della forza per la velocità. La massima potenza si sviluppa per un carico di circa il 30% della Vmax La regolazione del calcio mioplasmatico nel muscolo scheletrico REGOLAZIONE DELLA CONTRAZIONE SCOSSA SEMPLICE: risposta meccanica ad una singola stimolazione. Contrazione seguita da rilasciamento, con sviluppo di forza (o accorciamento) ridotto; la risposta meccanica dura almeno 10 volte più a lungo del potenziale d'azione. Scossa semplice REGOLAZIONE DELLA CONTRAZIONE TETANO INCOMPLETO: risposta meccanica a stimolazioni ripetute con intervalli più brevi del ciclo del Ca++ : rilasciamento incompleto e maggiore sviluppo di forza. Tetano incompleto REGOLAZIONE DELLA CONTRAZIONE TETANO COMPLETO (FUSO): risposta meccanica alla stimolazione ad alta frequenza (20-60 Hz). Non avviene rilasciamento e lo sviluppo di forza (o l'accorciamento) è massimo Tetano completo SPIEGAZIONI: progressivo accumulo* di Ca++ intracellulare all'avvicinarsi degli stimoli. Mantenimento in tensione degli elementi non contrattili, che sottraggono la maggior parte del lavoro compiuto dai sarcomeri nella scossa semplice. NB * la concentrazione degli ioni calcio nel tetano completo raggiunge e mantiene per più tempo il suo valore massimo che è lo stesso della scossa semplice o del tetano incompleto. Frequenza di stimolazione La concentrazione di Calcio raggiunge SEMPRE gli stessi valori La forza della contrazione può essere graduata dalla stimolazione ripetitiva che mantiene un’elevata concentrazione del calcio nel mioplasma durante il tetano completo ed incompleto. I singoli stimoli che provocano singoli potenziali d’azione sono indicati dalle linee verticali alla base della figura Vie metaboliche nel muscolo • Fosforilazione diretta • Glicolisi • Fosforilazione ossidativa Affaticamento di un muscolo scheletrico • Fatica cellulare (fatica neuromuscolare) • Fatica generale: condizione di omeostasi alterata provocata dal lavoro fisico
Scaricare