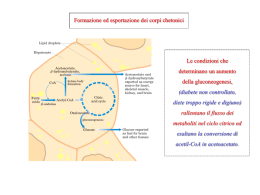
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 26 La gluconeogenesi Concetti chiave: • Il fegato e il rene possono sintetizzare glucosio da lattato, piruvato e amminoacidi. • La gluconeogenesi è per larga parte l'inverso della glicolisi, in cui la reazione della piruvato chinasi è sostituita dalle reazioni della piruvato carbossilasi e della fosfoenolpiruvato carbossichinasi. Le reazioni della fosfofruttochinasi e dell'esochinasi sono sostituite da reazioni catalizzate da fosfatasi. • La glicolisi e la gluconeogenesi sono reciprocamente regolate tramite effetti allosterici, fosforilazioni e cambiamenti della velocità di sintesi degli enzimi. La gluconeogenesi La gluconeogenesi è la sintesi di glucosio da precursori non glucidici. La riserva di glucosio dell’organismo (i.e. glucosio + glicogeno) è sufficiente per circa un giorno. Il mantenimento dei livelli di glucosio è importante dal momento che è il principale combustibile per il cervello (120g/die sui 160/die richiesti dall’intero organismo). La gluconeogenesi La gluconeogenesi ha luogo principalmente nel fegato, e in piccola quantità anche nel rene con la funzione di mantenere i livelli ematici di glucosio sufficientemente alti da consentire all’encefalo e al tessuto muscolare di avere sufficienti quantità di glucosio per soddisfare le proprie esigenze metaboliche. La gluconeogenesi che avviene nell’encefalo, nel muscolo scheletrico e nel muscolo cardiaco è solo di modesta entità. La gluconeogenesi La via gluconeogenetica converte il piruvato in glucosio. I precursori non glucidici del glucosio vengono prima convertiti in piruvato o entrano nella via a livello di intermedi successivi come l’ossalacetato e il diidrossiacetone fosfato. I principali precursori non glucidici sono il lattato, gli amminoacidi e il glicerolo. La gluconeogenesi La gluconeogenesi non è l’inverso della glicolisi. Alcune reazioni devono essere diverse perché in condizioni cellulari normali l’equilibrio è spostato verso la formazione del piruvato e non verso la sua conversione in glucosio. Tre reazioni della glicolisi sono così esoergoniche da essere praticamente irreversibili e sono quelle catalizzate dalla esochinasi, dalla PFK-1 e dalla piruvato chinasi. Nella gluconeogenesi queste tre tappe sono scavalcate. Glicolisi e gluconeogenesi a confronto Glucosio-6-fosfato Glicolisi e gluconeogenesi a confronto Conversione del piruvato a PEP Il fosfoenolpiruvato si forma dal piruvato in due reazioni che prevedono l’utilizzo di due gruppi fosforici ad alta energia: 1) Il piruvato viene prima carbossilato ad ossalacetato a spese di una molecola di ATP ad opera della piruvato carbossilasi (reazione anaplerotica del ciclo di Krebs) nel mitocondrio. 2) l’ossalacetato viene poi decarbossilato e fosforilato a spese di un secondo legame fosforico ad alta energia (GTP) ad opera della fosfoenolpiruvato carbossichinasi. La reazione complessiva è: Piruvato + ATP + GTP + H2O → Fosfoenolpiruvato + ADP + GDP + Pi + 2H+ Struttura a domini della piruvato carbossilasi La piruvato carbossilasi ha un dominio di legame dell’ATP che attiva lo ione HCO3- e trasferisce la CO2 al dominio di legame alla biotina (omologo al dominio della acetil CoA carbossilasi). Da qui la CO2 viene trasferita al piruvato nel dominio centrale. Il gruppo prostetico del carbossibiotinil-enzima La biotina è un gruppo prostetico legato covalentemente al gruppo ε-amminico di un residuo di Lys dell’enzima, che serve da trasportatore di CO2 attivato. Struttura a domini della piruvato carbossilasi La biotina si trova in una specie di braccio flessibile che le permette di spostarsi tra il sito specifico per l’ATPbicarbonato e il sito specifico per il piruvato. La carbossilazione del piruvato si svolge in tre stadi: 1. HCO3- + ATP → Carbossifosfato + ADP 2. Enzima-biotina + Carbossifosfato → CO2-enzima-biotina + Pi 3. CO2-enzima-biotina + piruvato → enzima-biotina + ossalacetato Meccanismo di reazione della piruvato carbossilasi Struttura a domini della piruvato carbossilasi La piruvato carbossilasi è un enzima mitocondriale mentre gli altri enzimi della via gluconeogenetica sono citosolici. L’ossalacetato prodotto dalla carbossilazione del piruvato viene trasportato nel citosol per generare fosfoenolpiruvato sottoforma di malato. Con il malato viene trasportato anche NADH. Il trasporto del PEP e dell'OAA Il PEP è trasportato direttamente tra citosol e mitocondrio. L’ossalacetato viene prima trasformato in aspartato dalla aspartato amminotransferasi (via 1) o in malato dalla malato deidrogenasi (via 2). La via 2 richiede l’ossidazione mitocondriale del NADH seguita dalla riduzione del NAD+ citosolico In tal modo vengono trasferiti equivalenti riducenti del NADH dal mitocondrio al citosol. Meccanismo di reazione della PEPCK L’ossalacetato viene simultaneamente decarbossilato e fosforilato dalla fosfoenolpiruvato carbossichinasi. Con la decarbossilazione viene favorita la reazione di fosforilazione altrimenti altamente esoergonica. Glicolisi e gluconeogenesi a confronto Una volta formatosi, il PEP viene metabolizzato dagli enzimi della glicolisi ma nella direzione inversa. Nelle condizioni intracellulari queste reazioni si trovano vicine all’equilibrio e quindi possono procedere fino alla successiva reazione irreversibile che è l’idrolisi del fruttosio 1,6-bisfosfato a fruttosio 6-fosfato e Pi. Glicolisi e gluconeogenesi a confronto Glucosio-6-fosfato Defosforilazione del F1,6-BP e sintesi di glucosio Il fruttosio 6-fosfato si forma dal fruttosio 1,6-bisfosfato per idrolisi dell’estere fosforico sull’atomo di carbonio C-1 ad opera della fruttosio 1,6 bisfosfatasi. La reazione complessiva è: Fruttosio 1,6-bisfosfato + H2O → fruttosio 6-fosfato + Pi Il glucosio libero si forma infine per idrolisi del glucosio 6-fosfato ad opera della glucosio 6fosfatasi presente nel fegato ma non nel muscolo e nel cervello. La reazione complessiva è: Glucosio 6-fosfato + H2O → glucosio + Pi Glucosio-6-fosfato Sintesi di glucosio dal glucosio-6-fosfato Nella maggior parte dei tessuti il glucosio 6-fosfato viene metabolizzato come tale, in particolare per formare glicogeno. Il vantaggio è che in tale forma non può uscire dalla cellula. La formazione del glucosio libero che avviene prevalentemente nel fegato ed in minor misura nel rene. Il glucosio 6-fosfato viene trasportato nel lume del reticolo endoplasmatico dove viene idrolizzato a glucosio dalla glucosio 6-fosfatasi, enzima legato alla membrana. Il glucosio e il Pi vengono poi trasferiti nel citosol ad opera di una coppia di trasportatori. Panoramica della gluconeogenesi Resa energetica della gluconeogenesi La reazione netta della formazione del glucosio dal piruvato è: 2 Piruvato + 4 ATP + 2 GTP + 2 NADH + 4 H2O → Glucosio + 4 ADP + 2 GDP + 6 Pi + 2 NAD+ + 2 H+ Per sintetizzare una molecola di glucosio dal piruvato vengono usati 6 legami fosforici ad alta energia e 2 molecole di NADH. Nella glicolisi durante la conversione del glucosio a piruvato vengono prodotte solo 2 molecole di ATP. Il costo aggiuntivo della gluconeogenesi è di quattro molecole ad elevato potenziale di trasferimento del gruppo fosforico per ogni molecola di glucosio sintetizzata dal piruvato. Tale costo è il prezzo da pagare per trasformare un processo energeticamente sfavorevole (inverso della glicolisi) in uno favorevole (gluconeogenesi). Ciclo di Cori L’alanina ed il lattato prodotti dal muscolo scheletrico in attività e dagli eritrociti (privi di mitocondri) sono uno dei principali rifornimenti della gluconeogenesi. La formazione del lattato nel muscolo sposta parte del carico metabolico dal muscolo al fegato. Il muscolo che si contrae fornisce lattato al fegato che lo utilizza per sintetizzare e rilasciare il glucosio. Il fegato ripristina dunque i livelli di glucosio necessari per le cellule muscolari attive che generano ATP attraverso la conversione glicolitica del glucosio in lattato (Ciclo di Cori). Alanina amminotrasferasi (ALT) L’alanina, insieme al lattato, è il più importante precursore del glucosio nel fegato quando, nel muscolo, gli scheletri carboniosi di alcuni amminoacidi vengono utilizzati come combustibili. ALT Gli atomi di azoto vengono trasferiti al piruvato formando alanina. La reazione inversa avviene nel fegato. Tale processo contribuisce a mantenere il bilancio azotato. Regolazione della gluconeogenesi Gluconeogenesi e glicolisi sono regolate reciprocamente e coordinati in modo tale che nella cellula le due vie non siano attive contemporaneamente. Alta carica energetica Bassa carica energetica Gluconeogenesi Glicolisi La logica della regolazione è la seguente: quando c’è bisogno di energia predomina la glicolisi; quando vi è un surplus di energia disponibile ha il sopravvento la gluconeogenesi. Regolazione della gluconeogenesi Il primo importante punto di controllo della gluconeogenesi è la reazione catalizzata dalla fruttosio 1,6bisfosfatasi. L’enzima glicolitico corrispondente è la fosfofruttochinasi-1. Alti livelli di ATP e citrato (alta carica energetica) inibiscono la fosfofrutto chinasi, il citrato attiva la fosfatasi. La regolazione ormonale (glucagone) della glicolisi e della gluconeogenesi nel fegato è mediata dal fruttosio 2,6 bisfosfato (F-2,6-BP). Il F-2,6-BP attiva la fosfofruttochinasi 1 e stimola la glicolisi nel fegato. Il F-2,6-BP inibisce la fruttosio 1,6 bisfosfatasi e rallenta la gluconeogenesi nel fegato. Regolazione della gluconeogenesi La concentrazione cellulare del modulatore fruttosio 2,6-bisfosfato viene determinata dalla velocità della sua sintesi da parte della fosfofruttochinasi-2 (PFK-2) e dalla velocità della sua degradazione operata dalla fruttosio 2,6-bisfosfatasi (FBPase-2). Regolazione della gluconeogenesi La fosfofruttochinasi-2 (PFK-2) e la fruttosio 2,6-bisfosfatasi fanno parte della stessa catena polipeptidica (enzima tandem) e sono regolati in modo complementare e coordinato dal glucagone. Regolazione della sintesi e degradazione del fruttosio 2,6-bisfosfato Una diminuzione della concentrazione del glucosio ematico, segnalato dal glucagone, determina la fosforilazione dell’enzima bifunzionale (aumento attività fosfatasica) e quindi fa diminuire la concentrazione di fruttosio 2,6-bisfosfato, rallentando la glicolisi. Regolazione della sintesi e degradazione del fruttosio 2,6-bisfosfato Un aumento della concentrazione di fruttosio 6-fosfato, che segue ad un aumento del glucosio ematico, accelera la formazione di fruttosio 2,6-bisfosfato facilitando la defosforilazione dell’enzima (aumento dell’attività chinasica). Regolazione della gluconeogenesi Glucosio abbondante ↑ Fruttosio 2,6 bisfosfato ↑ Glicolisi ↓ Gluconeogenesi Digiuno prolungato ↓ Fruttosio 2,6 bisfosfato ↓ Glicolisi ↑ Gluconeogenesi La gluconeogenesi è favorita quando la cellula è ricca di precursori biosintetici. Regolazione della gluconeogenesi Regolazione della gluconeogenesi Il secondo punto di controllo della gluconeogenesi è la reazione catalizzata dalla piruvato carbossilasi che è inibita dall’ADP ed è attivata dall’acetil CoA che, come il citrato, indica che il ciclo di Krebs sta producendo energia ed intermedi metabolici. Anche la fosfoenol-piruvato carbossichinasi è inibita dall’ADP. I cicli del substrato nel metabolismo del glucosio Un incremento del 20% della velocità di conversione di A in B (da 100 a 120) ed un decremento del 20% della velocità di conversione da B in A (da 90 a 72). Il nuovo flusso netto passa da 10 a 48 con un aumento del flusso netto del 380% a fronte di una variazione del 20%. I cicli del substrato amplificano i segnali metabolici.
Scaricare