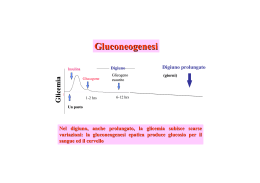
La gluconeogenesi è la via principale dei tessuti animali che porta alla formazione di carboidrati da precursori non saccaridici. Questa via è universale sia per gli animali, che le piante e i microrganismi; tuttavia, anche se le reazioni sono le stesse il contesto metabolico e le regolazioni della via differiscono da organismo a organismo e da tessuto a tessuto. Negli animali i precursori del glucosio sono il lattato, il piruvato, il glicerolo ed alcuni aminoacidi(glucogenici). Considerando che il cervello dell'uomo consuma circa 120 g. di glucosio al giorno e rappresenta la principale fonte d'energia per i tessuti embrionali, per gli spermatozoi, per la midollare del surrene e per i globuli rossi, si capisce quanto sia importante, nei tessuti animali, la biosintesi di glucosio. La gluconeogenesi avviene principalmente nel fegato dei mammiferi ed è citosolica, come la glicolisi, dalla quale differisce per tre deviazioni nelle reazioni irreversibili. Se la glicolisi converte il glucosio in piruvato, la gluconeogenesi converte il piruvato in glucosio e le due vie sono ovviamente soggette ad una regolazione coordinata e reciproca. Delle dieci tappe che trasformano il glucosio in piruvato nella glicolisi, sette fanno parte, in senso inverso, della gluconeogenesi. Le altre tre sono irreversibili, per cui vengono superate dalla gluconeogenesi con un pool di enzimi differente. Il fatto che questa sintesi passi per il mitocondrio non è casuale. Infatti l’uscita del malato dal mitocondrio porta fuori anche una molecola di NADH che scarseggia nel citosol ed è necessario per proseguire la sintesi di glucosio. La via alternativa preferita quando il precursore è il malato è più breve, infatti la conversione del lattato in piruvato genera NADH nel citosol. Per questo non è necessario che esca il malato dai mitocondri, ma può essere esportato direttamente il PEP. REAZIONI Il piruvato, dopo essere stato trasportato nei mitocondri viene convertito in ossalacetato dalla piruvato carbossilasi. La piruvato carbossilasi è il primo enzima regolatore della via, infatti l’acetil-CoA è un modulatore positivo dell’enzima. Questa reazione rappresenta una delle vie di ingresso nel ciclo dell’acido citrico. L’ossalacetato viene convertito in malato per essere trasportato al di fuori del mitocondrio dal trasportatore malato-a-chetoglutarato. Una volta uscito viene riconvertito in ossalacetato che a sua volta viene trasformato in fosfoenolpiruvato. Questa prima serie di reazioni richiede due gruppi fosforici ad alta energia (dall’ATP e dal GTP), mentre nella tappa inversa della glicolisi viene prodotta una sola molecola di ATP nella trasformazione da PEP a piruvato. Se queste due reazione potessero avvenire contemporaneamente si creerebbe un ciclo futile con dispersione di energia sotto forma di calore. Un ciclo futile analogo si potrebbe cerare anche a livello delle altre due deviazioni tra la glicolisi e la gluconeogenesi. Normalmente questi cicli sono impediti dalle regolazioni ma in certi casi possono diventare importanti per la produzione di calore. La prima deviazione dalla glicolisi avviene nella conversione da piruvato a fosfoenolpiruvato. Questa avviene secondo una via principale ad opera di una serie di enzimi sia citosolici che mitocondriali. Esiste anche una seconda via che è maggiormente attiva quando il lattato è precursore della gluconeogenesi. Il piruvato, dopo essere stato trasportato nei mitocondri viene convertito in ossalacetato dalla piruvato carbossilasi. Il piruvato può provenire anche dalla alanina per transaminazione (GPT). Esso, nei mitocondri, viene carbossilato dalla piruvico carbossilasi in ossalacetato (enzima già visto nelle vie anapletorica nel ciclo di Krebs). Tale enzima è il primo enzima regolatore della gluconeogenesi La seconda deviazione della gluconeogenesi rispetto alla glicolisi è la defosforilazione del fruttosio-1,6-bisfosfato in fruttosio-6fosfato. Questa reazione è catalizzata dall’enzima fruttosio -1,6bisfosfatasi. Questo enzima è regolato in maniera negativa dall’AMP e dal fruttosio-2,6-bisfosfato. La terza deviazione dalla glicolisi è la reazione finale della gluconeogenesi: la defosforilazione del glucosio-6-fosfato ad opera della glucosio-6-fosfatasi. Questo enzima è presente solamente negli epatociti, per cui la gluconeogenesi non può avvenire nel muscolo o nel cervello. Il glucosio prodotto nel fegato o ingerito con la dieta viene trasportato a questi tessuti dal flusso sanguineo. REAZIONE FINALE 2 Pyr+ 4 ATP + 2 GTP + 2 NADH + 4 H2O Glucosio + 4 ADP + 2 GDP + 6 Pi + 2 NAD + 2 H+ + La somma delle reazioni che portano alla formazione del di glucosio da piruvato comprende il consumo di sei legami fosforici ad alta energia e due molecole di NADH. La glicolisi invece produce solamente due molecole di ATP e due di NADH. Quindi la gluconeogenesi risulta una via relativamente costosa dal punto di vista energetico. Gran parte di questa energia è spesa per rendere le reazioni irreversibili diminuendo la variazione di energia libera. Si è accennato che alcuni aminoacidi possano essere utilizzati per la produzione di glucosio. Tra questi l’alanina e la glutammina sono particolarmente importanti perché trasportano gruppi amminici da tessuti extraepatici al fegato. Dopo il distacco dei loro gruppi amminici lo scheletro carbonioso è incanalato nella gluconeogenesi. Al contrario gli acidi grassi a catena pari non possono essere convertiti in glucosio in quanto dalla loro degradazione si ottiene solo acetil-CoA che a sua volta non può essere convertito in piruvato o ossalacetato. Tuttavia la degradazione degli acidi grassi è ugualmente importante per la gluconeogenesi perché produce, nei lunghi periodi di digiuno, gran parte dell’ATP e del NADH che servono alla gluconeogenesi. Per assicurarsi che i cicli futili di cui abbiamo parlato non avvengano in condizioni normali, la gluconeogenesi e la glicolisi devono essere regolate in maniera separata e complementare. ALTRI PRECURSORI GLUCOGENICI oltre a piruvato e lattato GLICEROLO Proprionil-CoA L’Acetil-CoA (e quindi i grassi) non è un precursore gluconeogenico REGOLAZIONE DELLA GLUCONEOGENESI REGOLAZIONE DELLA GLUCONEOGENESI La regolazione da parte dell’acetil-CoA dell’enzima piruvato carbossilasi ha come effetto di attivare la gluconeogenesi solo nel caso in cui la produzione di acetil-CoA supera la velocità di smaltimento nel ciclo dell’acido citrico. Quando le richieste energetiche della cellula sono soddisfatte rallenta la fosforilazione ossidativa con conseguente accumulo di NADH che inibisce il ciclo dell’acido citrico. L’accumulo di acetil-CoA inibisce la sua formazione dal piruvato attraverso la piruvato deidrogenasi e attiva la gluconeogenesi. Il secondo punto di controllo della gluconeogenesi è a livello dell’enzima fruttosio-1,6-bisfosfatasi, che è inibito dall’AMP. Il corrispondente enzima della glicolisi è invece attivato dall’AMP e inibito dall’ATP. Questo enzima subisce anche una seconda regolazione a livello ormonale. Il fegato ha anche la funzione di mantenere costante il livello di glucosio nel sangue. Quando il livello di glucosio diminuisce l’ormone glucagone segnala al fegato di produrre e rilasciare più glucosio. Una delle fonti di questo glucosio è la gluconeogenesi. La regolazione ormonale avviene attraverso il fruttosio-2,6-bisfosfato, effettore allosterico negativo della fruttosio-1,6-bisfosfatasi e positivo del corrispondente enzima della glicolisi. Questo modulatore è efficace a concentrazioni molto basse, per cui la sua sintesi risulta poco dispendiosa. La concentrazione del fruttosio-2,6-bisfosfato dipende dalla sua velocità di formazione e demolizione. Queste reazioni sono controllate dal glucagone attraverso il cAMP. Il glucagone stimola l’adenilato ciclasi e il cAMP stimola una proteina chinasi che fosforila l’enzima che produce il fruttosio-2,6-bisfosfato. La fosforilazione diminuisce la concentrazione del fruttosio-2,6-bisfosfato attivando la gluconeogenesi. Anche l’insulina è in grado di intervenire sulla gluconeogenesi. L’insulina è il segnale di abbondanza di sostanze nutrienti e stimola la conservazione dei combustibili e la sintesi di proteine in diversi modi. Sul fegato ha effetti opposti al glucagone e inibisce la gluconeogenesi. Infine va ricordato che anche i glucocorticoidi possono intervenire sulla gluconeogenesi agendo sulla trascrizione di alcuni enzimi coinvolti nella via biosintetica.
Scaricare