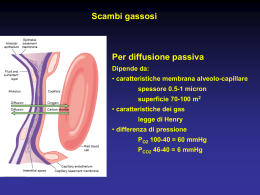
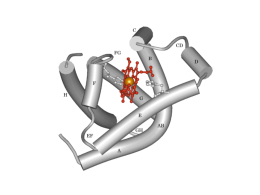
Scambio e trasporto di O2 e di CO2 Leggi dei gas: legge di Boyle • PV = nRT • In cui P è la pressione del gas (Atm), V è il volume che lo contiene (lt), n è il numero di molecole del gas, R è la costante universale dei gas, T è la temperatura assoluta • Da cui: • P = nRT/V • Poiché, data una certa quantità di gas in un contenitore, ad una certa temperatura n e T sono costanti, la P sarà inversamente proporzionale al V del contenitore, cioè: • P = 1/V Ciò è dovuto al fatto che le molecole di un gas sono in costante agitazione termica e, sbattendo contro le pareti del contenitore, esse esercitano su di esse una forza: la forza totale esercitata dalla collisione di tutte le molecole sulle pareti darà la P del gas. Quindi, se n è costante, ma il V diminuisce, aumenterà la densità delle molecole e quindi il numero di collisioni e, di conseguenza, la P. E viceversa. Inoltre, se, a parità di V, aumenta n, di nuovo aumenterà il numero di collisioni e quindi la P. Leggi dei gas: Legge di Dalton • La P esercitata da una miscela di gas è pari alla somma delle pressioni esercitate dai singoli gas che occupano lo stesso volume • La pressione esercitata dai singoli gas di una miscela viene chiamata pressione parziale di quel gas • Quindi la Ptot esercitata da una miscela di gas è pari alla somma delle pressioni parziali esercitate da ciascun gas che compone la miscela: • Ptot = P1 + P2 + P3 + P4 +….Pn • Dove n è il numero di gas che costituisce la miscela Pressioni parziali dei gas nell’aria atmosferica a 0°C e 760 mmHg (in condizioni di secchezza) GAS % Patm mmHg Pgas O2 20.95 di 760 = 159 PO2 CO2 0.04 di 760 = 0.3 PCO2 N2 79,01 di 760 = 596 PN2 Variando la temperatura ambiente e la percentuale di vapore acqueo si avranno valori diversi delle P parziali Pressioni parziali dei gas nell’aria alveolare (satura di vapore acqueo e a 37°C) A 37°C il vapore acqueo ha una P = 47mmHg, quindi se la P totale dei gas negli alveoli è 760 mmHg, O2, CO2 e N2 eserciteranno una P = 760 - 47 = 713 mmHg. Ma le percentuali dei tre gas nell’aria alveolare saranno diverse rispetto a quelle dell’aria atmosferica perché il sangue attraversando i capillari polmonari assume continuamente O2, diminuendone la concentrazione negli alveoli e cede CO2 aumentandone la concentrazione negli alveoli. Vale a dire che, nell’aria alveolare, le percentuali dei gas saranno rispettivamente di circa 15% per l’O2, 5% per la CO2 e 80% per l’N2. Quindi i valori delle P parziali dei gas saranno circa pari a: PO2 = 100 mmHg PCO2 = 40 mmHg PN2 = 573 mmHg Pressioni parziali nei liquidi • I gas mantengono gran parte delle loro caratteristiche molecolari quando si trovano in soluzione • Quindi anche nel sangue si possono valutare le pressioni parziali dell’O2 e della CO2 (N2 non partecipa agli scambi respiratori) • Se la pressione totale deve rimanere costante, quando aumenta una pressione parziale, l’altra deve ridursi Leggi dei gas: Legge di Henry • La solubilità di un gas in un liquido dipende dalla T, dalla P parziale del gas nella miscela gassosa all’interfaccia con il liquido, dalle proprietà chimiche del gas e dalle proprietà chimiche del liquido. Quindi, a T = K • c = KP • In cui c = concentrazione del gas disciolto, P è la pressione parziale del gas e K è la costante ad una data temperatura • Quindi a T = K la concentrazione di un gas in un liquido è proporzionale alla sua pressione parziale • Inoltre, se un gas si trova sopra un liquido, tale gas si scioglierà nel liquido finché non viene raggiunto l’equilibrio • In condizioni di equilibrio, le concentrazioni del gas nell’aria e nel liquido non saranno le stesse. Infatti, la concentrazione del gas nel liquido dipende dalla solubilità specifica di quel gas in quel particolare liquido: più è solubile, maggiore è la concentrazione del gas nel liquido • Quindi, quando aumenta la pressione parziale del gas aumenta anche la sua concentrazione nel liquido • Ma per gas diversi, a parità di pressione parziale, la concentrazione nel liquido sarà diversa in base alla diversa solubilità di ciascun gas nel liquido • Così, la concentrazione di CO2 nel plasma, a parità di pressione parziale, sarà superiore a quella della O2, perché superiore è la sua solubilità La diffusione semplice di una sostanza attraverso una membrana obbedisce alla legge di Fick Fattori che regolano la velocità di diffusione di O2 e di CO2 tra polmoni e sangue e tra sangue e cellule • La diffusione dei due gas attraverso le membrane obbedisce alla legge di Fick • Se si assume che la resistenza che offre la membrana alla migrazione dei due gas sia = K, la diffusione dei polmoni sarà direttamente proporzionale a: – 1) Gradiente di concentrazione (pressione parziale) – 2) Area della superficie di scambio • e sarà inversamente proporzionale a: – 3) Spessore della membrana – 4) Distanza di diffusione (la diffusione è più rapida su brevi distanze) • Poiché 2, 3 e 4 sono da considerarsi costanti, il fattore che più influisce sugli scambi è il gradiente di concentrazione Meccanismo degli scambi gassosi alveolari • Gli scambi gassosi sono dovuti a gradienti di pressione parziale dei singoli gas fra l’aria alveolare e il sangue • I gas si spostano dal luogo dove hanno pressione maggiore a quello dove la loro pressione è minore fino a raggiungere l’equilibrio • In particolare, nel polmone: l’O2 abbandona l’aria alveolare dove ha una PO2 di 100 mmHg e passa nel sangue polmonare dove la PO2 è di 40 mmHg; la CO2, invece, passa dal sangue polmonare, dove ha una P di 46 mmHg all’aria alveolare, dove ha una P di 40 mmHg • Viceversa, a livello tissutale: l’O2 abbandona il sangue dove ha una PO2 di 100 mmHg e passa nelle cellule dove la PO2 è di 40 mmHg; la CO2, invece, passa dalle cellule dei tessuti, dove ha una PCO2 di 46 mmHg, al sangue dove ha una PCO2 di 40 mmHg Gradiente pressorio e solubilità • Nel caso dell’O2 il gradiente pressorio necessario perché avvengano gli scambi gassosi (sia a livello polmonare, sia a livello tissutale) è di 60 mmHg • Nel caso della CO2 il gradiente pressorio necessario perché avvengano gli scambi gassosi (sia a livello polmonare, sia a livello tissutale) è di soli 6 mmHg • Ciò è dovuto al fatto che la solubilità della CO2 è 25 volte maggiore di quella dell’O2: pertanto lo scambio gassoso sarà sempre più veloce per la CO2 che non per l’O2 e quindi, per ottenere lo spostamento del gas, sarà sufficiente un gradiente pressorio di pochi mmHg Area della superficie di scambio • La ramificazione sequenziale dell’albero arterioso polmonare culmina con la formazione di una densa rete di capillari che avvolge gli alveoli • Questa rete alveolo-capillare ha un’area di superficie di circa 70 m2 (circa le dimensioni di un campo da tennis) Spessore della membrana e distanza di diffusione • Lo spessore della membrana è dato dalla matrice e dalle componenti tissutali della rete alveolo-capillare • Questa barriera comprende: le cellule epiteliali dell’alveolo di I tipo le cellule endoteliali dei capillari le loro rispettive membrane basali (che sono a contatto tra loro) • La distanza per lo scambio dei gas attraverso questa barriera è di circa 1-2 μm A = alveolo; c = capillare; col = collagene; el = elastina; e = endotelio capillare con nucleo (nu); I = epitelio di I tipo; * = interstizio Trasporto dei gas nel sangue • Poiché i gas non sono molto solubili nel plasma, oltre al loro dissolvimento, devono esistere dei sistemi di trasporto più efficienti • Infatti, ad es., a PO2 pari a 100 mmHg, il sangue contiene solo 3 ml di O2 per litro. Poiché, a riposo, i tessuti necessitano di 250 ml di O2/min, per sopperire alle necessità tissutali, la gittata cardiaca dovrebbe essere di circa 83 l/min anziché di 5 l/min come è in realtà • Quindi è chiaro che esistono altri sistemi di trasporto dei gas che garantiscono ai tessuti l’apporto necessario • I meccanismi di trasporto devono essere reversibili, in modo da consentire di legare l’O2 a livello polmonare e rilasciarlo a livello tissutale, e viceversa per quanto riguarda la CO2 Trasporto dell’ossigeno • Ciascun litro di sangue arterioso contiene circa 200 ml di ossigeno: – 3 ml disciolti nel plasma o nel citosol degli eritrociti (1,5%). Solo questo ossigeno disciolto contribuisce alla PO2 sanguigna – 197 ml (98,5%) trasportati legati all’emoglobina, proteina che si trova negli eritrociti L’emoglobina Cromoproteina costituita da 4 subunità proteiche, due α e due β. Ciascuna subunità contiene una globina (catena polipeptidica globulare) fissata con un legame covalente ad un gruppo eme (protoporfirina) che contiene Fe++ Concentrazione dell’emoglobina nel sangue • Varia con l’età: nel neonato è pari a circa 20 g/100 ml di sangue, nell’adulto ha un valore di circa 15 g/100 ml nel maschio e 14g/100 ml nella femmina • Si ha anemia quando la concentrazione di emoglobina scende sotto i 13 g/100 ml nel maschio e sotto i 12 g/100 ml nella femmina • In alcune situazioni, come l’adattamento ad alta quota, si può avere un aumento dei globuli rossi (policitemia) e della concentrazione di emoglobina per assicurare un adeguato apporto di ossigeno ai tessuti anche in presenza di una diminuita PO2 Emoglobina e ossigeno • Ciascun gruppo eme della emoglobina è in grado di legare una molecola di ossigeno • Quindi ogni molecola di emoglobina può trasportare fino a 4 molecole di ossigeno • Ogni legame di una molecola di O2 con l’Hb aumenta l’affinità dell’Hb per l’ O2 e quindi aumenta la probabilità che altro O2 si leghi (cooperazione positiva) • Il complesso costituito da emoglobina e ossigeno è detto ossiemoglobina (HbO2) • Una molecola di emoglobina priva di ossigeno è detta desossiemoglobina Nei polmoni • L’ossigeno si sposta dall’aria alveolare al sangue capillare e si lega all’emoglobina Nei tessuti Quando il sangue raggiunge i tessuti, le molecole di ossigeno si dissociano dall’emoglobina e diffondono alle cellule Reversibilità di legame • Affinché l’emoglobina sia utile al trasporto di ossigeno, è necessario che essa si leghi all’ossigeno in maniera reversibile • Il legame deve quindi essere sufficientemente forte da poter legare grandi quantità di ossigeno a livello polmonare, ma non così forte da non poterlo cedere ai tessuti Il legame emoglobina-O2 dipende dalla PO2 • Il legame o il rilascio di ossigeno dipende dalla PO2 del sangue: – un’elevata PO2 facilita il legame dell’O2 con l’emoglobina – una bassa PO2, ne facilita il distacco Saturazione dell’emoglobina Nel sangue arterioso la PO2 è di 100 mmHg e l’emoglobina è saturata al 98,5%, cioè quasi tutti i siti di legame sono occupati dall’O2. Pochissimo ossigeno è disciolto nel citosol degli eritrociti Saturazione dell’emoglobina Nel sangue venoso misto la PO2 è di 40 mmHg e l’emoglobina è saturata al 75%, cioè ogni 4 siti di legame, 3 sono occupati dall’ossigeno Curva di dissociazione dell’emoglobina • A pressioni parziali basse, poco O2 si lega all’Hb. • All’aumentare della PO2 la quantità di O2 che si lega all’Hb prima aumenta rapidamente, poi tende a stabilizzarsi quando la saturazione si avvicina al 100%. • Come mostra la tabella, alle normali pressioni parziali nelle arterie e nelle vene sistemiche, a riposo, la percentuale di saturazione dell’Hb varia solo del 25% circa. Variazioni dell’affinità dell’Hb per l’O2 • Esistono per lo meno altri quattro fattori (T, pH, 2,3-DPG, PCO2) che possono influenzare l’affinità dell’Hb per l’O2 • Tali cambiamenti si riflettono in uno spostamento verso destra o verso sinistra della curva di dissociazione dell’Hb • Lo spostamento della curva verso destra indica una diminuzione di affinità, cioè indica che l’O2 viene liberato più facilmente dall’Hb, rendendolo disponibile ai tessuti • Viceversa, uno spostamento della curva verso sinistra indica che l’O2 viene caricato più facilmente dall’Hb per via di una maggiore affinità • Quindi, per uno spostamento verso destra è necessaria una PO2 più alta di 45mmHg per ottenere una saturazione dell’80%, per uno spostamento verso sinistra è sufficiente una PO2 inferiore ai 45mmHg per determinare la stessa percentuale di saturazione Effetti della modificazione dell’affinità dell’Hb per l’O2 sulla curva di dissociazione Il legame emoglobina-O2 è influenzato dalla Temperatura La T influenza l’affinità dell’Hb per l’O2 alterando la struttura dell’Hb (come fa per la struttura terziaria di tutte le proteine, quindi effetto non specifico) Poiché all’aumentare del metabolismo la T aumenta, diminuendo l’affinità dell’Hb per l’O2, l’O2 viene rilasciato più facilmente ai tessuti, soprattutto se molto attivi Viceversa, la diminuzione della T del sangue durante il passaggio nei polmoni, promuove il legame tra Hb e O2 Il legame emoglobina-O2 è influenzato dal pH (effetto Bohr) Quando la concentrazione degli H+ aumenta, l’affinità dell’Hb per l’O2 diminuisce, viene rilasciato più O2 e la curva si sposta verso destra Ad es. nel corso del metabolismo anaerobio delle fibre muscolari si produce acido lattico che rilascia H+ nel citoplasma e nel liquido extracellulare con abbassamento del pH Il legame emoglobina-O2 è influenzato dalla PCO2 Nel sangue pH e PCO2 sono strettamente correlati: un aumento della PCO2 si accompagna ad una diminuzione del pH (con spostamento verso destra della curva e diminuzione dell’affinità per l’O2) e viceversa Ma le modificazioni della curva dovute ad una variazione della PCO2, non sono totalmente spiegabili sulla base della variazione del pH perché esiste anche un fattore specifico regolato esclusivamente dalla PCO2 Il legame emoglobina-O2 è influenzato dal 2,3-DPG (2,3-difosfoglicerato) • Il 2,3-DPG è prodotto dagli eritrociti come prodotto intermedio della glicolisi • L’ossiemoglobina ad alte concentrazioni inibisce l’enzima che forma il 2,3-DPG e quindi i livelli di 2,3-DPG sono bassi ed hanno poco effetto sull’affinità dell’Hb Ma se i livelli di ossiemoglobina sono bassi (come quando i rifornimenti di O2 sono ridotti), interviene la sintesi del 2,3-DPG che diminuisce l’affinità dell’Hb per l’O2 favorendone il rilascio nei tessuti che ne hanno bisogno (anemia, altitudine) Il legame emoglobina-O2 e il monossido di C Il monossido di carbonio (gas inodore, incolore prodotto nella combustione incompleta dei materiali organici) è tossico in quanto si lega più velocemente dell’O2 all’Hb formando carbossiemoglobina Esso impedisce il legame dell’O2 con l’Hb e ne diminuisce, di conseguenza, la capacità di trasportare O2 del sangue Il legame mioglobina-O2 • La mioglobina è il pigmento rosso del muscolo che ha una struttura simile a quella di una delle catene di emoglobina • Possiede un solo gruppo eme e può quindi legare una sola molecola di ossigeno La sua curva è spostata a sinistra rispetto a quella dell’Hb, quindi: l’O2 ha una maggiore affinità per la mioglobina, viene caricato più facilmente e ceduto ai tessuti con maggiore difficoltà rispetto all’Hb Trasporto della CO2 • Ciascun litro di sangue arterioso contiene circa 490 ml di anidride carbonica: – 27 ml disciolti nel plasma o nel citosol degli eritrociti (5,5%). – 24 ml (5%) trasportati legati all’emoglobina come carbaminoemoglobina – 439 ml (90%) disciolto nel sangue sotto forma di bicarbonato. Il bicarbonato viene formato a partire dalla CO2 negli eritrociti dei capillari sistemici (dove i livelli di CO2 sono più alti) grazie alla presenza dell’enzima anidrasi carbonica L’anidrasi carbonica Nell’eritrocita avviene la seguente reazione: CO2 + H2O anidrasi carbonica H2CO3 H+ + HCO3- Un aumento di CO2 (o di PCO2) sposta la reazione verso destra (producendo più ioni idrogeno e bicarbonato) Una diminuzione della CO2 (o della PCO2) sposta la reazione a sinistra, (producendo più anidride carbonica a partire da ioni H+ e HCO3-) Scambio della CO2 a livello tissutale • Respirando, a riposo le cellule producono circa 200 ml/min di CO2 che diffonde nell’interstizio e poi nel plasma, grazie al gradiente pressorio • La CO2 nel plasma aumenta e quindi aumenta la PCO2 che risulterà maggiore della PCO2 degli eritrociti • Quindi la CO2 diffonde dal plasma agli eritrociti per gradiente pressorio e solo una piccola parte resta disciolta Negli eritrociti una parte si lega all’emoglobina e la maggior parte viene trasformata in ioni bicarbonato e idrogeno grazie all’anidrasi carbonica In tal modo la PCO2 del plasma diminuisce in modo da mantenere un gradiente che consenta alla CO2 di continuare a spostarsi dai tessuti al plasma Anche la CO2 negli eritrociti, trasformata in ioni HCO3-, diminuisce: quindi essa può continuare a spostarsi dal plasma agli eritrociti • Se gli ioni H+ e HCO3- si accumulassero negli eritrociti, la reazione tra anidride carbonica e acqua potrebbe raggiungere un equilibrio • In tal caso, la CO2 non verrebbe più convertita in ioni idrogeno e bicarbonato e si accumulerebbe nelle cellule in forma disciolta determinando un aumento della PCO2 dentro gli eritrociti • Il sangue, quindi, non potrebbe più trasportare CO2 sotto forma di bicarbonato • Per impedire questo, è necessario che gli ioni idrogeno e bicarbonato vengano rimossi dagli eritrociti • Quando il livello degli ioni bicarbonato negli eritrociti aumenta, essi vengono trasportati nel plasma e scambiati con gli ioni Cl grazie ad una proteina di trasporto che si trova nella membrana plasmatica degli eritrociti (scambio dei cloruri) • Gli ioni idrogeno vengono tamponati dall’emoglobina, con formazione di emoglobina ridotta (HHb) • Il legame degli idrogenioni all’Hb è alla base dello spostamento a destra della curva di saturazione dell’Hb con cessione di O2 ai tessuti da parte dell’Hb Riassumendo: Carbaminoemoglobina (piccola quantità) Scambio della CO2 a livello polmonare A livello alveolare, la CO2 passa dal plasma agli alveoli, per gradiente di concentrazione, per essere espirata La perdita di CO2 fa sì che gli ioni HCO3- ed H+ negli eritrociti, si ricombinino ad H2CO3 che, per effetto dell’anidrasi carbonica, viene riconvertito in H2O e CO2 che diffonde negli alveoli per essere espirata Man mano che gli ioni HCO3- degli eritrociti vengono utilizzati, i loro livelli dentro gli eritrociti diminuiscono e nuovi ioni HCO3- entrano dal plasma in scambio con il ClLa CO2 che diffonde negli alveoli per essere espirata, favorisce la formazione di altra CO2 Riassumendo: Curva di saturazione della CO2 • Così come la PCO2 del sangue influenza l’affinità dell’Hb per l’O2, anche la PO2 influenza l’affinità dell’Hb per la CO2 • In particolare, un aumento della PO2 determina uno spostamento a destra della curva, diminuendo l’affinità dell’Hb per la CO2 e quindi la sua capacità di trasportare CO2 (effetto Haldane) La diminuita affinità dell’Hb per la CO2 (effetto Haldane) è dovuta al fatto che, quando la PO2 aumenta, l’Hb trasporta più O2 e si riduce la quantità di CO2 che può legarsi all’Hb Riassunto del trasporto dei gas Nei tessuti Nei polmoni
Scaricare