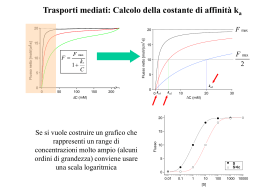
Attività elettrica dei neuroni Neuroni piramidali del SNC che stanno crescendo su una matrice di glia/fibroblasti Un esperimento in current-clamp Si inietta una corrente I di intensità nota nella cellula e si misura la conseguente variazione del potenziale di membrana Vm Quindi, ad ogni valore della corrente iniettata possiamo associare un determinato valore di Vm Riportiamo in un grafico i valori ottenuti: 6 6 5 5 Corrente (nA) Corrente (nA) Riportiamo adesso sullo stesso grafico altri valori ottenuti da un’altra cellula: 4 3 4 3 2 2 1 1 -70 -60 -50 -40 -30 -20 -10 0 10 Pot. di Membrana (mV) La relazione corrente/voltaggio è lineare, come c’è da aspettarsi se vale la legge di Ohm: I=g·Vm dove g = costante Qual è la differenza? -70 -68 -66 -64 -62 -60 Pot. di Membrana (mV) In questo caso la relazione corrente/voltaggio non è lineare, quindi non vale la legge di Ohm: I=g·Vm cioè g ≠ costante Alcuni tipi di canali Ionici sono sempre aperti, altri no! extracell. intracell. Sempre aperti Il loro comportamento è ohmico: la relazione I/V è lineare g=costante Voltaggio-dipendenti: rispondono a variazioni di Vm Il loro comportamento non è ohmico: la relazione I/V non è lineare perché g non è costante [g=f(Vm)] Riassumendo Se sulla membrana esistono canali voltaggio-dipendenti, le variazioni del potenziale di membrana Vm in risposta ad uno stimolo elettrico dipendono: oltre che dal tempo necessario a caricare la membrana, che si comporta come un condensatore (proprietà passive di membrana), anche dal fatto che gm varia al variare del voltaggio e del tempo, gm=f(V,t) (proprietà attive dei canali). Il metodo di indagine del current-clamp non permette di separare la componente resistiva della corrente (IR), dovuta alla presenza di canali ionici, da quella capacitiva (IC), dovuta al comportamento della membrana come un condensatore La derivazione delle correnti ioniche transmembranarie amplificatore Neurone piramidale con un elettrodo di vetro attaccato per la derivazione delle correnti ioniche transmembranarie mediante la tecnica del (patch-) voltage-clamp La tecnica del voltage-clamp consiste nel bloccare il potenziale di membrana ad un valore costante nel tempo e nel registrare le correnti ioniche transmembranarie generate a tale potenziale di membrana Potenziale di membrana finale (Vm) costante (mV) 20 Potenziale di riposo (Vr) -70 0 10 20 Tempo (ms) 30 40 Corrente ionica (nA) 60 Corrente transmembranaria generata dal passaggio di Vm da –70 mV a +20 mV 40 20 0 -20 -40 -60 0 10 20 30 40 Tempo (ms) Vantaggi del voltage-clamp: In genere, gm = f(V,t) ma, 1. Vm è bloccato ad un valore costante gm = f(t) soltanto e può essere dedotto dall’andamento della corrente ionica Ii 2. E’ possibile separare Ii da Ic. Infatti cm è caricata istantaneamente Voltaggio (mV) Separazione farmacologica delle correnti IK e INa 20 -70 0 10 20 Tempo (ms) 30 40 + TTX (IK) INa+IK Corrente ionica (nA) 60 40 20 0 + TEA (INa) -20 -40 -60 0 10 20 Tempo (ms) 30 40 Voltaggio-dipendenza dei canali Kv 140 120 100 + I(K ) max (nA) Voltaggio (mV) 60 50 40 30 20 10 0 -10 -20 -30 80 60 40 IK=gK• (V-EK) 20 0 -70 0 10 20 30 -20 40 -30 -20 -10 0 Tempo (ms) 10 20 30 40 50 60 Voltaggio (mV) 140 1.0 120 Conduttanza (μS) Corrente K + (nA) 100 80 60 40 0.8 0.6 0.4 gK = IK/(V-EK) 20 0.2 0 0.0 -20 0 10 20 Tempo (ms) 30 40 EK=-80 mV -30 -20 -10 0 10 20 30 40 50 60 Voltaggio (mV) Quesito del giorno Un ricercatore, in seguito ad esperimenti condotti in voltageclamp ha ricavato che la corrente di K+ (IK) dipende dal potenziale di membrana (V) secondo la seguente relazione: 1 IK 4.6 (V 85) 1 (V 10) / 9 1 e (pA) 1) Calcolare i valori di IK nell’intervallo di potenziali tra –80 e +80 mV ad intervalli di 10 mV e metterli in grafico in funzione di V; 2) Dai valori di IK ricavati, e sapendo che EK= -85 mV, calcolare i valori corrispondenti di gK e metterli in grafico in funzione di V; 3) A quale valore di V sara’ aperta la meta’ dei canali? IK = 4.6·[1-1/(1+EXP((V+10/9))]·(V+85) (pA) EK = -85 mV IK 0.0 0.1 0.4 1.9 7.1 24.7 74.1 172.5 294.2 394.3 466.4 522.9 572.8 620.2 666.7 712.9 759.0 Vm -EK 5 15 25 35 45 55 65 75 85 95 105 115 125 135 145 155 165 gK 0.002 0.006 0.018 0.053 0.158 0.450 1.139 2.300 3.461 4.150 4.442 4.547 4.582 4.594 4.598 4.599 4.600 600 400 200 0 -100 -50 0 50 100 mV nS Vm -80 -70 -60 -50 -40 -30 -20 -10 0 10 20 30 40 50 60 70 80 pA 800 5.0 4.0 3.0 2.0 1.0 0.0 -100 -50 0 mV 50 100 Voltaggio-dipendenza dei canali Na 1 0 40 30 20 10 0 -10 -20 -30 -40 -50 -1 I(Na) picco (nA) Voltaggio (mV) 60 -2 -3 -4 -6 -70 -7 0 10 20 30 40 -50 -40 -30 -20 -10 0 10 20 30 40 0 -1 -2 -3 -4 -5 0.12 gNa = INa/(VENa) 0.10 ENa=+50 mV 0.14 0 10 20 Tempo (ms) 30 40 0.08 0.06 0.04 0.02 0.00 -50 -40 -30 -20 -10 0 10 20 30 40 0 10 20 Tempo (ms) 60 Voltaggio (mV) Conduttanza (μS) I(Na) (nA) I(Na) (nA) Tempo (ms) 0 -1 -2 -3 -4 -5 -6 -7 INa=gNa• (V-ENa) -5 30 40 Voltaggio (mV) A parità di conduttanza, è la driving force che determina l'andamento della relazione I/V EA =+55mV Vm -80 -60 -40 -20 0 20 40 60 80 g (nS) 0.00 0.04 0.89 4.71 5.56 5.60 5.60 5.60 5.60 Vm -EA -135 -115 -95 -75 -55 -35 -15 5 25 EB=-80mV IA (pA) -0.2 -4.3 -84.5 -353.3 -305.9 -196.0 -84.0 28.0 140.0 Vm -EB 5 25 45 65 85 105 125 145 165 1000 800 EA=+55mV EB=-80mV Corrente (pA) 600 400 200 0 -200 -400 -100 -80 -60 -40 -20 0 20 Voltaggio (mV) 40 60 80 100 IB (pA) 0.0 0.9 40.0 306.2 472.8 587.9 700.0 812.0 924.0 Quesito del giorno Un ricercatore, in seguito ad esperimenti condotti in voltageclamp e utilizzando la tossina di scorpione per bloccare l’inattivazione, ha ricavato che la conduttanza del canale del Na+ (gNa) dipende dal potenziale di membrana (V) secondo la seguente relazione: 1 gNa 5.6 1 (V 30) / 6 (nS) 1 e 1) Calcolare i valori di gNa nell’intervallo di potenziali tra – 80 e +80 mV ad intervalli di 10 mV e metterli in grafico in funzione di V; 2) Dai valori di gNa ricavati, e sapendo che ENa= +55 mV, calcolare i valori corrispondenti di INa e metterli in grafico in funzione di V. (nS) ENa=+55 mV nS gNa=5.6(1-1/[1+EXP((V+30)/6)]) 6.0 5.0 4.0 gNa (nS) 0.00 0.01 0.04 0.19 0.89 2.80 4.71 5.41 5.56 5.59 5.60 5.60 5.60 5.60 5.60 5.60 5.60 Vm -ENa -135 -125 -115 -105 -95 -85 -75 -65 -55 -45 -35 -25 -15 -5 5 15 25 INa (pA) -0.2 -0.9 -4.3 -20.3 -84.5 -238.0 -353.3 -351.5 -305.9 -251.7 -196.0 -140.0 -84.0 -28.0 28.0 84.0 140.0 3.0 2.0 1.0 0.0 -100 -50 0 50 100 50 100 mV pA Vm -80 -70 -60 -50 -40 -30 -20 -10 0 10 20 30 40 50 60 70 80 200 100 0 -100 -50 -100 0 -200 -300 -400 mV Canali voltaggio-dipendenti - CINETICA Il movimento termico fa oscillare la proteina-canale tra la conformazione aperta e quella chiusa Quindi, questa transizione Chiuso Aperto è un evento casuale La legge delle probabilità ci permette tuttavia di fare certe predizioni sul comportamento medio del canale Come esempio consideriamo un canale che ha soltanto uno stato chiuso e uno stato aperto. a Closed b u.d.m: s-1 Open Calcolo della Probabilità di apertura (Po) dPO/dt = aPC – bPO PO + PC = 1 dPC/dt = bPO - aPC PC = 1 - PO dPO/dt = a – aPO – bPO dPO/dt = a – (a + b)PO Risolvendo l’equazione differenziale e ponendo PO(t)=n(t), si ottiene: n(t) = a/(a b)a/(a b)exp[(a b)t]a0/(a0 b0)exp[(a b)t] In particolare, all'equilibrio: dPO/dt = a - (a b)PO = 0 PO eq = a/(a b) = n Ponendo inoltre a0/(a0 b0) = n0 e 1/(a b) = t si ottiene: n(t) = n - (n - n0) exp(t/t) Essa rappresenta la relazione tra le costanti di velocità e, • a livello microscopico: la probabilità che un singolo canale con un’unica gate sia aperto, • a livello macroscopico: la frazione di canali (unica gate) aperti 60 -70 160 140 120 100 80 60 40 20 0 -20 Prob. di apertura (Po) I(K) Voltaggio (mV) Cinetica delle correnti di K+ del canale Kv 1.0 0 10 20 Tempo (ms) 30 40 Ipotesi: una sola gate Po=n(t)=n∞-(n∞-n0)∙exp(-t/τ) I=g∙(V-E) ; gPo I(t)n(t)∙(V-E) I=g∙(V-E) ; gPo=n4 0 10 20 Tempo (ms) 30 I(t)n4(t)∙(V-E) chiuso aperto -70mV +60mV 40 Ipotesi: quattro gates identiche Po= n∙n∙n∙n = n4 n4(t)=[n∞-(n∞-n0)∙exp(-t/τ)]4 1 gate 0.8 4 gates 0.6 0.4 0.2 0.0 0 10 20 Tempo (ms) 30 40 chiuso aperto Volt. (mV) Cinetica delle correnti di Na+ voltaggio-dipendenti 20 -70 0 10 20 30 40 Ipotesi: tre gates identiche Po=m3(t)=[m∞-(m∞-m0)∙exp(-t/τ)]3 Tempo (ms) 0 I(Na) -5 -10 -15 Chiuso Aperto -70mV +20mV -20 -25 0 10 20 30 40 Prob. di apertura (Po) Tempo (ms) m(t) 1.0 Ipotesi: tre gates identiche di attivazione + una gate di inattivazione Po=m3(t)∙h(t) h(t) 0.8 m3(t) 0.6 0.4 0.2 m3*h 0.0 0 10 20 Tempo (ms) 30 40 Chiuso Aperto Inattivato -70mV +20mV +20mV Quesito del giorno Depolarizzando un neurone dal potenziale di riposo Vo ad un certo potenziale Vf=+10 mV, la probabilita’ di apertura di una singola gate di attivazione “n” del canale del K+ varia nel tempo seguendo la seguente relazione temporale: n(t) = 0.8·[1- EXP(-t / 1.1) ], dove il tempo t e’ espresso in ms. Sapendo che quel tipo di canale del K+ ha 4 gates “n” identiche e che la conduttanza massima GK e’ 25 nS: 1. calcolare e mettere in grafico i valori della conduttanza gK(t) ad intervalli di 1 ms per una durata totale di 12 ms; 2. calcolare il valore di IK allo stato stazionario (EK = -80 mV). n(t) = 0.8·[1- EXP(-t / 1.1) ] EK=-80 mV GK=25 mS g=G·n4 IK= g·(V-EK) = G·n4·(V-EK) t(ms) 0 1 2 3 4 5 6 7 8 9 10 11 12 n 0.000 0.478 0.670 0.748 0.779 0.792 0.797 0.799 0.799 0.800 0.800 0.800 0.800 4 n 0.000 0.052 0.202 0.313 0.368 0.392 0.403 0.407 0.408 0.409 0.409 0.410 0.410 g (nS) 0.00 1.30 5.04 7.81 9.20 9.81 10.07 10.17 10.21 10.23 10.24 10.24 10.24 IK (pA) 0.0 117.2 453.8 703.2 828.2 883.1 905.9 915.3 919.0 920.6 921.2 921.4 921.5 12 10 8 6 4 2 0 0 2 4 6 8 10 12 14 Ancora un problema Il potenziale di riposo di una cellula è determinato dalla presenza di due canali permeabili rispettivamente agli ioni A e B attraverso i quali passano le correnti IA e IB indicate in tabella. Dopo aver disegnato i rispettivi grafici I/V, determinare: 1) le conduttanze gA e gB; 2) se i due canali sono voltaggio-dipendenti o -indipendenti; 3) il potenziale di equilibrio dei due ioni A e B; 4) il potenziale di riposo Vr della cellula. V(mV) -120 -110 -100 -90 -80 -70 -60 -50 -40 -30 IA (nA) -1650 -1100 -550 0 550 1100 1650 2200 2750 3300 V(mV) -20 -10 0 10 20 30 40 50 60 70 IB (nA) -1320 -1100 -880 -660 -440 -220 0 220 440 660 V(mV) -120 -110 -100 -90 -80 -70 -60 -50 -40 -30 IA =gA (V-EA ) (nA) -1650 -1100 -550 0 550 1100 1650 2200 2750 3300 V(mV) -20 -10 0 10 20 30 40 50 60 70 IB=gB(V-EB) (nA) -1320 -1100 -880 -660 -440 -220 0 220 440 660 4000 3000 2000 EA EB 1000 0 -150 -100 -50 0 -1000 -2000 EA=-90mV g1A=I[(-120)-I(-110)]/[-120-(-110)]= 55 nS EB=+40mV g1B=I[(-20)-I(-10)]/[0-(-10)]= 22 nS Vr=(EAgA+EBgB)/(gA+gB)= -52.9 mV 50 100 Confronto della voltaggio-dipendenza di gK e gNa 1.0 Conduttanza g 0.8 0.6 0.4 0.2 0.0 -60 -40 -20 0 Vm (mV) 20 40 60
Scaricare