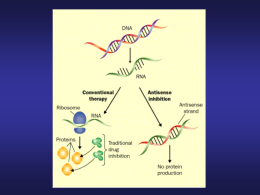
CORSO DI LAUREA SPECIALISTICA IN BIOTECNOLOGIE DEL FARMACO Adriana Maggi BIOTECNOLOGIE FARMACOLOGICHE AA 2008/09 LEZIONE 6 Acidi nucleici e farmaci GLI OLIGONUCLEOTIDI ANTI-GENE (TRIPLEX) L’oligonucleotide si lega in modo specifco al DNA, intercalandosi nella struttura a doppia elica, formando un tratto di catena tripla che determina l’inattivazione della replicazione del DNA e della trascrizione dell’mRNA VANTAGGI DELLA TERAPIA ANTIGENE RISPETTO AGLI ODN Questa strategia comporta la formazione di un breve tratto a tripla elica (mediante legami di Hoogsteen) nelle regioni di controllo della trascrizione presenti nel DNA. Tale modalità di impiego mostra prospettive alquanto interessanti poiché l’azione diretta sul gene implica l’uso di quantità inferiori di ODN essendo in questo caso solo due i targets (le due copie del gene) rispetto ai numerosissimi targets della strategia ODN (le copie dell’mRNA). Si garantisce inoltre una più durevole attività terapeutica contando sulla maggiore durata dell’emivita del gene rispetto a quella del messaggero. TRIPLEX Le prime indicazioni, indirette circa la possibilità di formazione di un tratto di DNA a tripla elica risale a studi sulla densità ottica con miscele di poli U e poli A condotti da B. Felsenfeld nel 1957. Hoogsteen chiarisce la natura dei legami che consentono la formazione della tripla elica in tratti omopurinici e omopirimidinici (AG o TC). In questo contesto la C non presenta atomi disponibili per un ponte idrogeno con la G prospiciente e deve essere quindi protonata. Tale reazione può avvenire a un pH leggermente acido (6,6) e in presenza di ioni bivalenti. In queste condizioni l’ODN ibridizza con la sequenza omopurinica complementare mediante legami di Hoogsteen, formando così le due triplette T-AT e C-GC. L’ODN si colloca nel solco maggiore del duplex preesistente con una orientazione parallela alla sequenza omopurinica del DNA bersaglio. LEGAMI DI HOOGSTEEN PROBLEMATICHE DELLA TERAPIA ANTI-GENE 1. Il bersaglio deve essere omopuriniche/pirimidiniche costituito da sequenze 2. La lunghezza della sequenza bersaglio deve essere minimo di 15 nucleotidi consecutivi 3. Il rapporto A/G non deve essere inferiore a 4 molte sequenze 5’ rispetto ai DNA codificanti hanno tratti omopurinici 4. Tutte le C devono essere protonate e quindi occorre agire a pH più bassi di quello fisiologico. Questo problema può essere superato con alcuni stratagemmi METODICHE PER SUPERARE L’OSTACOLO DELLA PROTONAZIONE 1. utilizzo nella sintesi dell’ODN di 5-metilcitosina che consente l’ibridizzazione al duplex anche a pH neutri 2. Utilizzare ODN costituiti da PNA che hanno una migliore capacità di ibridizzare 3. Utilizzo di molecole intercalanti legate all’ODN, capaci di legare una base sul duplex conferendo una maggiore stabilità alla tripla elica 4. Sostituire la base citosina con l’eterociclo 2aminopiridina, il cui pKa è maggiore di quello della citosina e ciò favorisce il legame a pH neutro CHIMICA DEL LEGAME PNA-DNA I PNA possono legarsi al DNA sia attraverso legami idrogeno sia di Hoogsteen CH3 5’ 5’ 3’ CH3 3’ T 3’ 3’ O N 5’ 5’ N O O H H N H N PNA Strand invasion Hoogsteen T H N N N N O N Watson - Crick A Watson-Crick + NH 3 PNA Strand C T C T T C T T C DNA Strand '3 G A G A A G A A G 5' C T C T T C T T C NH3+ Hoogsteen PNA Strand COO - COO- I RIBOZIMI RIBOZIMI NATURALI Producono nel taglio di un RNA substrato estremità 3’OH e 5’ fosfato. Sono raggruppabili in classi in base alla loro struttura ed alle reazioni in cui sono coinvolti RNAsi P: Ubiquitaria, è responsabile della maturazione dei tRNA nel loro estremo 5’ RNAsi HRP: Replicazione DNA mitocondriale tagliando RNA primer INTRONI tipo I: Tetrahymena pre RNA mitocondriale di lievito geni di cloroplasti INTRONI tipo II: con meccanismo di splicing conservato (precursori di mRNA in organelli di funghi e piante) CARATTERISTICHE COMUNI AI RIBOZIMI Reazione di formazione e rottura di legami covalenti in molecole substrato a RNA I ribozimi sono catalizzatori di reazioni che li vedono altresì implicati come substrato La struttura tridimensionale dell’enzima determina la sua specificità di legami con il substrato a creare il sito catalitico Le reazioni catalizzate sono basate sullo scambio di protoni (con conseguente idrolisi o transesterificazione risultanti nel taglio di legami fosfodiesterici) I ribozimi sono metallo-enzimi MECCANISMO D’AZIONE DEGLI INTRONI DI TIPO I 1. Attacco nucleofilo della guanina presente nell’ambiente 2. Generazione di un terminale OH 3. Seconda transesterificazio ne per attacco nucleofilo del terminale libero sul primo nucleotide dell’esone in 3’ SITI ATTIVI NEGLI INTRONI DI TIPO I ex2 3' 3'G414 OH 414 G G-binding site G-OH 5' G ex1 5' Substrate binding site 3' CUCUCU GGGAGG 2nd transfer IGS UUUACCU GGGAGG ex1 + ex2 1st transfer ex2 3rd transfer G414 ex1 5' G 414 CUCUCU OH GGGAGG G 5' GGGAGG + 5' G UUUACCU MECCANISMO D’AZIONE DEGLI INTRONI DI TIPO II • Usano il 2’ OH di una adenina interna all’introne 2’ A • Attacca il 5’ fosfato sul Exon 1 HO Exon 2 primo nucleotide dell’introne • L’introne assume una struttura circolare tipica detta “lariat” 2’ A • Gli esoni sono uniti e l’introne OH è exciso mantenendo la struttura “lariat” • Un enzima appositamente Exon 1+2 deputato taglia il “lariat” e 2’ A + origina un introne lineare • L’introne lineare viene degradato STRUTTURA DEI RIBOZIMI HAMMERHEAD Lo schema rappresenta il cambiamento di struttura in due fasi, indotto dal legame di ioni Mg2+. La prima fase consiste nella formazione di un dominio coassiale tra due eliche la seconda genera il core catalitico. Il ribozima con la struttura finale è in grado di auto-tagliarsi nel punto indicato dalla freccia rossa STRUTTURA SECONDARIA E TERZIARIA DEI RIBOZIMI HAMMERHEAD La sequenza CUGA adiacente allo stem loop I è sufficiente per il taglio Struttura terziaria STRUTTURA CRISTALLOGRAFICA DEI RIBOZIMI HAMMERHEAD La struttura cristallina evidenzia 5 siti di legame per il Mg2+ PICCOLI RNA CATALITICI-1 VIROIDI Molecole di RNA infettive che funzionano indipendentemente, prive di capside proteico VIRUSOIDI Simili ai viroidi ma sono incapsulati insieme al genoma virale in virus di piante. Non replicano indipendentemente dal virus. Sono perciò detti RNA satelliti PICCOLI RNA CATALITICI-MECCANISMO D’AZIONE 1. legame fosfodiestere intatto 2. Stato di transizione che coordina il mg2+ 3. Taglio del legame fosfodiestere Questi RNA promuovono reazioni di auto-taglio. Tale reazione è favorita da cationi divalenti e genera terminali 5’ OH e 2’-3’ ciclici. Molti di essi per tagliare assumono una struttura hammerhead RIBOZIMI MECCANISMO D’AZIONE -1 RNA è una molecola meno stabile del DNA perché l’O in 2’ può attaccare il legame fosfodiesterico adiacente (attacco nucleofilo) e romperlo, lasciando un fosfato 2’-3’ ciclico e un ossidrile in 5’. MODALITA’ DI TAGLIO DI RNA: RNAsi P e introni di tipo I e II catalizzano l’idrolisi del legame fosfodiesterico, lasciando un fosfato in 5’ e un ossidrile in 3’ Le reazioni di taglio che dipendono dall’O in 2’ possono essere attivate in due modi: 1) Ambiente alcalino: l’O in 2’ si attiva per ionizzazione 2) Ribonucleasi A catalizza la reazione di taglio usando un residuo basico di istidina per spostare il protone dell’O al 2’ e un residuo acido di aspartato per trasferirlo al 5’ RIBOZIMI MECCANISMO D’AZIONE -2 La carica positiva di un residuo di lisina adiacente interagisce con il fosfato e lo stabilizza durante lo stato di transizione. Il fosforo tende ad assumere una conformazione a bipiramide trigonale durante lo stato di transizione e questo può spiegare perché il taglio mediato dall’O al 2’ è più rapido in presenza di nucleotidi (pirimidina, adenosina) specifici perché le interazioni tra basi possono influenzare la conformazione del legame fosfodiestere APPLICAZIONI TERAPEUTICHE DEI RIBOZIMI-1 Sono attivi in “CIS” cioè intramolecolarmente. Studi in corso prevedono manipolazioni per indurli a lavorare in “TRANS” ad esempio per il riparo di RNA mutati o danneggiati. Esempio: ripristino del controllo della proliferazione cellulare agendo su p53 mRNA mutato mRNA corretto + Ribozima con mRNA corretto mRNA scartato + + ribozima RIBOZIMI IN CLINICAL TRIALS-1 ANGIOZYME TM Ribozima disegnato per inibire il VEGF (Vascular Endothelial Growth Factor). ANGIOGENESIS HEPTAZYME TM Ribozima che lega sequenze altamente conservate del virus dell’epatite C HCV DRUG RESISTANCE RIBOZIMI IN CLINICAL TRIALS-2 HIV Gene Therapy... Bone Marrow Sample Treat Stem Cells with Retroviral Vector Encodes Gene for anti-HIV Ribozyme Re-Implant Treated Cells APTAMERI Sono ORN o ODN capaci di riconoscere una specifica sequenza amminoacidica o un epitopo appartenente ad una proteina Sono stati scoperti per caso osservando l’alterazione del processo di coagulazione, dovuto ad un aumento del tempo di protrombina in seguito alla somministrazione di ODN antisenso. In queste molecole e’ presente un “G quartet” che conferisce al DNA la capacità di legare la trombina inattivandola APTAMERO PER L’a-TROMBINA Organizzazione strutturale dell’aptamero per l’a-trombina e disposizione dei quartetti di guanina che ne stabilizzano la conformazione APTAMERI CON STRUTTURA 3D DETERMINATA Acido nucleico Affinita’ Kd (mM) Teofillina RNA 0,3 FMN RNA 0,5 AMP DNA/RNA 6/10 Arginina DNA/RNA 125/60 Citrullina RNA 65 Tobramicina RNA 0,009 Neomicina B RNA 0,115 HIV-1 Rev peptide RNA 0,004 HTLV-1 Rex peptide RNA 0,025 MS2 coat protein RNA ND Trombina DNA 0,025 Ligando STRUTTURA DI ALCUNI APTAMERI Riconoscimento di ligandi aromatici piatti: A: strutture B: teofillina-RNA C: FMN-RNA D: AMP-DNA E: AMP-RNA Riconoscimento dell’antibiotico aminoglicoside tobramicina: A: struttura B: aptamero tobramicina-RNA C: la tasca che accoglie il ligando Riconoscimento di aminoacidi basici: A: strutture B,C: arginina-DNA D: arginina-RNA E: citrullina-RNA Riconoscimento di peptidi e proteine: A:peptide derivante dalla proteina Rev del virus umano HIV1 B,C: diversi aptameri RNA Rev D, E: aptamero proteina di rivestimento del batteriofago MS2RNA MOTIVI STRUTTURALI CHE RICONOSCONO IL DNA FLESSIBILITA’ DEL DNA Struttura del complesso CAP-DNA. Il DNA risulta ripiegato in due punti in seguito all’interazione con la proteina. I cilindri rappresentano la regione proteica interessata. SELEZIONE DEGLI APTAMERI-1 A tutt’oggi è impossibile indicare a priori se esistano e quali siano le sequenze di DNA in grado di riconoscere una definita struttura enzimatica L’analisi basata sui metodi computazionali richiede tempi di elaborazione esageratamente prolungati per stabilire la conformazione spaziale di macromolecole L’isolamento di aptameri ad attività biologica richiede lo sviluppo di metodologie sperimentali specifiche SELEX ( Systematic evolution of ligands by exponential amplification) Random or doped DNA pool Regenerated DNA pool Trascribed RNA pool cDNA 1. Generazione di una diversità di sequenza (sintesi chimica, manipolazione enzimatica, PCR) 2. Selezione di forme funzionali attraverso il legame con il bersaglio e l’eliminazione di molecole non affini 3. Amplificazione della popolazione selezionata con metodiche di PCR 4. Step successivi di selezione e riamplificazione per isolare le molecole più efficienti UTILIZZO DEGLI APTAMERI L’UTILIZZO TERAPEUTICO è ancora incerto Sono utili MEZZI DIAGNOSTICI grazie alla loro alta affinità e specificità per il bersaglio. Come gli anticorpi possono essere legati a molecole radioattive o ad enzimi la cui attività può essere facilmente valutata. Sono però più piccoli e maneggevoli degli anticorpi APTAMERI IN TERAPIA Il primo aptamero approvato come farmaco è stato il MACUGEN per il trattamento della degenarazione maculare (OSI Pharmaceuticals). Archemix (Cambridge, Ma) sviluppa aptameri antagonisti del Fattore di Willebrand attualmente in Fase clinica 2 per sindrome acuta coronarica Aptameri non modificati sono in studio per la coagulazione ematica e per la terapia di patologie oculari APTAMERI IN DIAGNOSTICA Aptamero legante della tenascina per imaging di neoplasie in sviluppo da parte della Schering AG SVANTAGGI DEGLI APTAMERI COME FARMACI 1. Come gli aODN sono molecole relativamente poco stabili 2. Problemi nella distribuzione e nella biodisponibilità 3. Costo di produzione NB. Possono però essere modificati chimicamente per aumentarne la stabilità e si possono utilizzare le metodologie di veicolazione già viste per gli ODN VANTAGGI DEGLI APTAMERI COME FARMACI 1. Interagiscono specificatamente con i loro target 2. La grande area superficiale degli acidi nucleici permette la formazione di più interazioni con il bersaglio rispetto ai farmaci classici, ciò determina un legame molto stretto (Kd nel range nanomolare e picomolare) 3. Sono in grado di inibire la funzione del loro bersaglio ma anche di modificare il metabolismo associato a quel bersaglio es. aptameri anti-trombina bloccano anche la coagulazione del sangue DECOY Sono brevi sequenze di DNA omologhe ad una sequenza consenso per una proteina transattivante. Il decoy legandosi inibisce in riconoscimento consenso modo della la funzione della competitivo al sito la sequenza proteina con proteina di MECCANISMO D’AZIONE DI DECOY CHE INIBISCONO LA PROLIFERAZIONE CELLULARE Ciclina A cdk P RB E2F Silente TTTCGCGC C-myc C-myb Cdc2 Kinase PCNA Ciclina A cdk RB P E2F TTTCGCGC Ciclina A cdk RB E2F TTTCGCGC P Transattivante TTTCGCGC TTTCGCGC TTTCGCGC TTTCGCGC Silente DECOYS IN THERAPY oligonucleotide mimicking a negative regulatory sequence of promoter C of estrogen receptor alpha (ER alpha) gene is sufficient for its re-expression in ER-negative human cancer cell lines NFκB decoy oligo therapy for IBD (Inflammatory Bowel Disease) E2F Decoy Transfection by HVJ-AVE–Liposome Method Cardiac Allograft Arteriopathy NUOVI TARGET TERAPEUTICI DIFFERENZE APTAMERI/DECOY APTAMERI • RNA o DNA filamento singolo o doppio • Omologhi di consensus dell’RNA • legano proteine citoplasmatiche DECOY • ODN a doppia elica • Omologhi di consensus dell’DNA • legano proteine nucleari
Scaricare