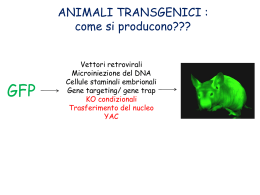
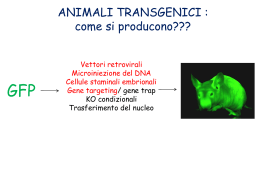
Organismi Transgenici - Alterazione del genoma tramite tecniche di manipolazione del DNA - Metodo diverso rispetto all’induzione di mutazioni - Le mutazioni indotte avvengono in maniera random - Prime prove di organismi transgenici : inserzione di un elemento esogeno nel genoma - Preparazione di costrutti adatti per essere attivi in genomi di origine diversa, Nei genomi eucariotici non esistono unità autonome autoreplicanti come i plasmidi nei batteri. Perché? Meiosi e mitosi vogliono strutture cromosomiche con centromero Topi transgenici per espressione e knock-out Inserzione random Ricombinaz. omologa (cellule ES) Embrioni tetraploidi Costrutti con BAC Mutanti condizionali Espressione inducibile Topi transgenici per inserzione random nel genoma Primi topi transgenici solo di espressione di marcatori, geni eterologhi, sovrannumerari, - iniezione diretta del DNA nella blastocisti - inserzione in una o più copie nel genoma ospite, - inizialmente un marcatore carrier per il colore del mantello Metodologia di base - organismo transgenico Come si ottiene: trasfezione batterica come modello Trasfettare cellule eucariotiche animali e vegetali col DNA Tecnica del DNA ricombinante per i vettori Cellule vegetali hanno anche la parete di cellulosa: metodi di trasfezione più complessi Cellule animali hanno solo la membrana meno resistente Tecniche di trasfezione diverse e con efficienze diverse Necessità della tecnica adeguata per veicolare il DNA Modelli transgenici Animali modello: drosofila, coenorabditis, xenopus, topo Essenziale è avere la finestra in cui intervenire nel processo di sviluppo dell’animale Nel topo si riesce ad isolare la blastocisti Tutto il lavoro preliminare era stato fatto dagli embriologi Cellule ES Blastocisti di topo Blastocisti al Microscopio elettronico Procedura per topi transgenici Da cosa dipende l’efficienza di espressione del transgene: - sito di inserzione -numero di copie che si inseriscono nel genoma - Il controllo dell’ integrazione con Southern o con PCR. - Con PCR inversa per trovare il punto in cui si è inserito. - L’espressione si verifica con la tecnica adatta per determinare il fenotipo. - Si usane anche geni reporter. Topi transgenici per knock-out - ricombinazione omologa in cellule ES (staminali embrionali totipotenti solo recenti) - la frequenza di ricombinazione è circa di 10-6. Le cellule staminali ES si coltivano in vitro e si deve evitare che differenzino. - vettore per ricombinazione omologa ha regioni di omologia al sito in cui deve ricombinare ed un gene per la selezione con un antibiotico - elettroporazione del vettore nelle cellule staminali. Vettori per ricombinazione omologa Struttura ad - costrutti ad con la parte esterna (le spalle) con l’omologia al gene “target” da mutagenizzare, - nella parte interna è presente un gene per la selezione con un antibiotico per cellule di mammifero (neomicina) ed anche un gene reporter ( gal, GFP ecc.). - L’inserzione provoca la mutagenesi del gene endogeno che se è letale non darà origine ad un topo transgenico. L’espressione dei geni per la resistenza ad antibiotici o dei geni reporter deve essere controllata da promotori costitutivi o inducibili per poter fare la selezione delle cellule ricombinanti. (prova con i geni Hprt e tk timidino-kinasi. I mutati danno la sensibilita’ al terreno HAT hypoxanthine, aminopterin and thymidine e solo Hprt produce resistenza alla 6-thioguanina). esperimenti di ricombinazione omologa di Capecchi Knock-out del gene per la resistenza HGPRT ipoxantina-guanina fosforibosil transferasi. Il problema e’ di selezionare le cellule con un fenotipo, ma sono pochi i geni con un fenotipo selettivo, in tale assenza spesso si utilizza la res. per un antibiotico. Il costrutto per far avvenire la ricombinazione omologa deve avere una regione omologa a quella con cui vogliamo ottenere la ricombinazione: mut. w.t. vettore a struttura - costrutto con due regioni di omologia (spalle) rispetto alla regione interna che si vuole inserire. La frequenza e’ stata studiata ed e’ ~ 1- 2 x10 -6 Per studiare la ricombinazione nel topo ci vorrebbero milioni di topi per 1 ricombinante invece in una fiaschetta si coltivano molti milioni di cellule: circa 106 / ml Mutanti condizionali Il sistema cre-lox Sfruttando il sistema di ricombinazione del fago P1 con i siti Lox e la ricombinasi Cre, si possono ottenere dei mutanti che perdono la regione voluta solo attivando la ricombinasi Cre. Si devono costruire dei vettori con la regione genetica da eliminare con siti Lox all’esterno (introni) e si chiamano floxed. Con questa strategia si possono ottenere topi transgenici per geni che sono letali in fasi diverse e soprattutto con l’espressione della ricombinasi Cre tessuto specifica, si può far avvenire il knock-out del gene solo in particolari tessuti dove si esprime o dove si induce Cre. Topi transgenici mutanti condizionali Il limite principale dei topi transgenici knock-out per un gene e’la possibile mortalita’ degli omozigoti durante lo sviluppo. - una soluzione per questo problema potrebbe essere l’induzione della mutazione tempo e tessuto specifica - e’ stato applicato il sistema di ricombinazione fagica (fago P1) Cre-Lox utilizzato anche in biologia cellulare perche’ inducibile - Cre e’ la ricombinasi (causa ricombinazione) e LoxP (locus di crossover in P1), serve per mantenere una sola copia del genoma, evita i multimeri - i siti Lox sono costituiti da palindrome di 13 bp ed una regione centrale di 8bp - la ricombinasi fa avvenire lo scambio di filamento tra due strutture Lox allineate originando delezione se sono in tandem, inversione se sono in palindrome, duplicazione se sono su cromatidi fratelli e integrazione se c’e’ un elemento in un plasmide ed un altro nella sequenza dove si integra (vedi fig. 1) inversion intramolecular deletion a b a c tandem a c palindrome c a b a b d c b c scambio su cromat. fratelli a a a c c b b c intramolecular deletion / duplication a b integration c - Questa tecnologia e’ applicata alle cellule staminali di topo ES. - L’inserzione dei siti LoxP si deve far avvenire tramite gene-targeting con un costrutto specifico (i costrutti con i geni con le regioni fiancheggianti che contengono siti LoxP si chiamano “floxed”) - quando si esprime il gene Cre avviene la ricombinazione e se si esprime solo in un tessuto la mutazione diventa tessuto specifica e si puo’ seguire il destino delle cellule che lo vanno a formare, per esempio nelle cellule pancreatiche quelle che formeranno le cell. a e - l’espressione del gene Cre non sempre e’ omogenea nel tessuto e la ricombinazione avviene a mosaico non nel 100% delle cellule - come controllo si usano due geni reporter, per cui se si ha ricombinazione il primo fiancheggiato da due siti Lox (lacZ) si inattiva e si attiva il secondo gene reporter (di solito la GFP o la fosfatasi alcalina) L’introduzione delle cassette per la resistenza ed i siti LoxP si fa avvenire per ricombinazione omologa Il costrutto per il gene floxed costrutto esone x induz. di Cre ricombinaz. e delez. neo esone y esone x introne x loxP - neo - loxP neo esone x introne x loxP esone z gene tk loxP loxP - neo - loxP ricombinaz. omologa costrutto ricomb. introne x esone z esone y gene tk loxP esone y esone z loxP gene tk Il gene per la timidino kinasi del virus Herpes simplex rende le cellule sensibili al Ganciclovir Nel caso di ricombinazione non omologa il gene tk non verra’ eliminato e le cellule potranno essere eliminate con la selezione dell’antibiotico Dopo l’eliminazione della resistenza alla neomicina per l’induzione del gene Cre le cellule ES mutanti sono pronte per essere iniettate nelle blasocisti Il gene per la ricombinasi Cre deve essere indotto e deve esprimersi al momento giusto e nel posto giusto Ci sono linee di topi transgenici che esprimono Cre nelle cellule epatiche, oppure nel sistema nervoso centrale SNC o altri sistemi Sono state fatte proteine di fusione con il dominio mutato di legame al ligando (ligand binding domain LBD) dell’ormone degli estrogeni, in modo che la ricombinasi sia confinata nel citoplasma finche’ non viene somministrato l’ormone sintetico che riconosce il recettore e fa traslocare la proteina di fusione con la ricombinasi nel nucleo dove induce la delezione del frammento incluso tra i due siti LoxP (vedi esempio della figura precedente) Analisi del sistema CRE-LOX per topi transgenici con “Gene Targeting” sito e tempo specifici. (Methods 24 , 2001, pag 71-80) D.Metzger e P. Chambon - Il Gene targeting ha alcune limitazioni per l’assenza della funzione del gene colpito da mutazione (targetet) che durante le fasi di sviluppo puo’ risultare letale precludendo lo studio di funzioni possibili negli stadi iniziali e successivi a quello letale. - Inoltre molti geni svolgono funzioni multiple in vari tipi di cellule durante l’ontogenesi e fase postnatale (pleiotropici), questo provoca fenotipi complessi e complica l’individuazione di cellule anomale prodotte da fenomeni con piu’ cause. - E ancora, l’effetto di una mutazione puo’ essere compensata durante lo sviluppo mascherando il fenotipo alterato nell’animale adulto. Nel caso di famiglie di geni si devono mutare piu’ geni della stessa famiglia per prevenire la ridondanza funzionale di espressione che preclude l’identificazione di una funzione di un componente di quella famiglia genica. - Definire una funzione di un gene membro di una famiglia puo’ essere ancora piu’ complicato quando la famiglia e’ coinvolta in un sistema pleiotropico di un pathway di segnali come i recettori per l’acido retinoico o FGF. - Altri effetti confondenti il knock-out di un gene possono essere il rischio di danno sulla fertilita’e disordini sistemici. - In tutti questi casi sara’ problematico determinare la funzione di un gene in una frazione di cellule ad un certo momento della vita del topo. - inoltre nel caso di modelli animali di patologie umane con mutazioni somatiche come il cancro - Quindi la necessita’ di avere un metodo con l’inattivazione condizionale di un gene e’ molto forte Sono state sviluppate strategie per avere gene-targeting condizionale in topo basato su cellule tessuto-specifico o espressione inducibile sito specifica del gene della ricombinasi CRE del fago P1. Ligand Inducible Cre Recombinase L’attivita’ di molti enzimi (oncoproteins, fattori di trascriz., RNA-binding prot., kinasi) puo’ essere controllata in maniera dipendente dal Ligando se fusa al dominio che lega il ligando LBD (ligand binding domain) di un recettore di un ormone steroideo. E’ stata fatta una proteina chimerica attiva della ricomb. Cre - LBD del recettore dell’estrogeno (ER) per cui l’attivita’ della ricombinasi Cre-ER dipende dalla presenza di 17-estradiol. Per non avere il controllo della proteina in presenza di estradiolo endogeno: e’ stato mutato il LBD del recettore ER (Cre-ERT) in maniera da legare solo un ligando sintetico (tamoxifen =Tam, 4idrossi tamoxifen=OHT) sfruttando la mutazione nota (Gly 525 Arg). Questo costrutto funziona in cellule in vitro. -Esperimento di espressione del gene in topo transgenico : costrutto messo sotto il controllo del promotore/enhancer del gene IE di CMV (citomegalovirus) il frammento PvuII del costrutto pCMVCre-ERT iniettato in zigoti F1(C57BL/6XSJL), analisi del transgene da DNA caudale. Analisi del topo transgenico Dopo l’analisi del DNA della coda per PCR Analisi dell’espressione con anticorpi anti CRE, scarsa espressione eccetto nell’epidermide. Analisi della presenza dell’ RNA estratto da vari tessuti Transgene Cre - ERT PvuII CMV promoter 3 globin intron SV40 polyA signal inizio trascrizione INTRON Cre-ERT 1 poly A 4 2 PvuII Prova di espressione e funzionamento su un topo transgenico “floxed” per il gene del recettore dell’acido retinoico a RXR 7 E8 E9 8 RXRa(targetet floxed gene) 5 Tk neo E8 LoxP RXRa(targetet floxed gene Excision of floxed marker After CRE recombinase) 50 40 30 20 Prodotto di PCR 156 bp Primers 7 e 8 7 E8 E9 8 E9 6 LoxP Prodotto di PCR 190 bp Primers 7 e 8 100 80 60 40 10 20 0 0 mRNA CRE Er % RXRa wt Livello di espressione di CRE (pallini) e di excisione dopo 3 giorni e dopo 1 giorno nella coda Metodo del gene trapping Per fare topi transgenici su vasta scala generalizzata, senza un solo target In realtà il topo transgenico si ottiene solo dopo aver selezionato la cellula staminale, quindi il vero prodotto è solo la collezione di cloni di cellule staminali ES mutagenizzate nei diversi geni. RNA interference (hairpin double strand RNA) Metodi per interferire con l’espressione di un gene: “stable suppression of gene expression by RNAi in mammalian cells” P.N.A.S. vol.99 n.3 pp 1443-8 P.J.Paddison et al. - mutagenesi, mutazioni termo sensibili(condizionali), soppressori - knock out per ricombinazione - anticorpi contro la proteina (prodotti da un vettore) - RNA antisenso trascritto da un vettore - oligo antisenso Questi metodi hanno il difetto di non essere sempre applicabili ai sitemi eucarioti; in certi casi non sono regolabili. L’RNA antisenso per funzionare deve essere aggiunto in dosi massicce, oppure deve essere trascritto dentro la cellula, ma il funzionamento della interferenza e’ legata al sistema Dicer, cioe’ ad un pathway enzimatico che riduce l’RNA specifico del messaggero in frammenti. E’ stato fatto un esperimento sfruttando il metodo CRE Lox per produrre un RNA a doppio filamento per bloccare l’enzima Dicer stesso Soppressione stabile dell’espressione genica tramite RNA interference in cellule di mammifero Sistema cellulare di difesa antivirale dimerizzazione PKR fosforilazione Blocco non specifico della traduzione EIF2a (kinasi) dsRNA esogeno (~ 500 bp) attiva Cofattore per la ribonucleasi non - specifica (Rnasi L) 2’- 5’ oligodenilato polimerasi pcDNA3 P GFP ZEO r L GFP L ZEO r Gene per la resistenza alla zeocina; L Lox P; GFP Le prime 500 bp codificanti di enhanced gr. fluor. prot. EGFP; P Promotore di citomegalovirus; P GFP ZEO r L GFP L Ricombinasi CRE P GFP ZEO r ZEO L GFP L r dsGFP Rapporto FF:REN Riduzione di 10 volte dell’espressione di FF in cellule trasfettate con i due plasmidi FF/REN FF = firefly luciferase REN = renilla luciferase ds = double strand ss = single strand as = antisense
Scaricare