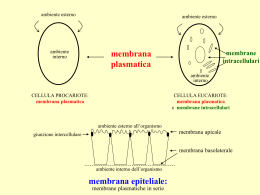
Biologia e sistematica vegetale Le membrane cellulari Le cellule richiedono le membrane per la loro esistenza… La membrana plasmatica è quella che definisce i confini della cellula e aiuta a creare e mantenere ambienti elettrochimicamente distinti tra l’interno e l’esterno della cellula. Le membrane cellulari: delimitano i contorni della cellula e dei suoi diversi compartimenti interni. Funzioni della membrana: 1. 2. 3. Foto al TEM di una sottile sezione criofissata di una cellula del meristema dell’apice radicale 4. 5. Delimitare contorni della cellula e dei suoi compartimenti Costituire siti di specifiche funzioni Contenere proteine di trasporto per regolare il movimento di sostanze tra interno ed esterno o tra i vari organelli Contenere i recettori necessari per rilevare i segnali esterni Fornire i dispositivi per la comunicazione cellula-cellula e per l’adesione cellulare. Queste barriere delimitano compartimenti nei quali la composizione chimica può differire da quella delle regioni circostanti ed essere ottimizzata per un’attività particolare. Le membrane possono inoltre fungere da “impalcatura” per alcune proteine. Rappresentazione schematica di una cellula del mesofillo fogliare, raffigurante i principali sistemi di membrane ed i domini di una parete cellulare di una cellula vegetale differenziata LA MEMBRANA PLASMATICA Controlla il trasporto delle molecole verso l’interno e l’esterno della membrana; Trasmette i segnali dall’ambiente alla cellula; Partecipa alla sintesi e all’assemblaggio delle molecole della parete cellulare; Fornisce legami fisici tra gli elementi del citoscheletro e la matrice extracellulare. Al pari di tutte le membrane biologiche,la membrana plasmatica mostra una permeabilità selettiva: permette, cioè, ad alcune sostanze di attraversarla più facilmente di altre. Esistono molte affinità tra membrana plasmatica vegetale e quella animale e le differenze sono in parte dovute alla contiguità con le pareti cellulari e ai rapporti con l’apoplasto. Le membrane Barriera di permeabilità. Permeabilità selettiva a determinate sostanze Divisione in compartimenti funzionali Es. Mitocondrio: contiene enzimi, substrati e le molecole utili per il processo della respirazione aerobica Lisosoma: contiene enzimi per la degradazione selettiva di proteine e lipidi. Lipidi e proteine si associano con interazioni non covalenti a costituire una pellicola di ~ 8nm di spessore. Modello di membrana a mosaico fluido Secondo il modello di SINGER & NICOLSON,oggi largamente accettato, le membrane sono delle strutture appiattite,tridimensionali, formate da proteine e lipidi. Si tratta di un mosaico di proteine affondate in un doppio strato lipidico fluido, i cui componenti sono in costante movimento. In questo modello alcune delle proteine della membrana si muovono, più o meno liberamente, nel doppio strato fluido. Poiché le proteine e le molecole dei lipidi si muovono lateralmente all’interno del doppio strato, le proteine formano arrangiamenti, o mosaici, che variano nel tempo e da sito a sito da cui il nome a “MOSAICO FLUIDO” ! Il modello a “mosaico fluido” illustra come i caratteri meccanici e fisiologici della membrana siano definiti dalle proprietà chimico - fisiche dei vari componenti cellulari. LE MEMBRANE DELLA CELLULA HANNO LA STESSA STRUTTURA DI BASE: Un Doppio Strato Lipidico E Proteine Globulari Le cellule vegetali contengono sistemi differenti di membrane. TIPI DI MEMBRANE PRESENTI NELLE CELLULE VEGETALI MEMBRANA PLASMATICA MEMBRANA NUCLEARE RETICOLO ENDOPLASMATICO CISTERNE DEL GOLGI (CIS, MEDIANO, TRANS) VESCICOLE DI TRASPORTO RIVESTITE DI CLATRINA MEMBRANA DELLE VESCICOLE DI ENDOCITOSI MEMBRANA ENDOSOMICA MEMBRANE DEL CORPO MULTIVESCICOLARE/VACUOLO AUTOFAGICO TONOPLASTO MEMBRANA DEL PEROSSISOMA MEMBRANA DEL GLIOSSISOMA MEMBRANE DEL CLOROPLASTO MEMBRANE DEI TILACOIDI MEMBRANE DEI MITOCONDRI Dal momento in cui si formano le cellule,per sopravvivere, devono mantenere l’integrità di tutti i compartimenti delimitati da membrane, cosicché tutti i sistemi di membrane devono passare da una generazione di cellule alla successiva in una forma funzionalmente attiva. L’eredità delle membrane segue alcune regole: le cellule figlie ereditano una serie completa di membrane dalla loro madre ciascuna cellula madre potenziale mantiene una serie completa di membrane le nuove membrane vengono prodotte soltanto mediante crescita e fissione di membrane esistenti; non possono essere costruite de novo Mentre il doppio strato fosfolipidico fornisce la struttura di base e la natura impermeabile delle membrane cellulari, le proteine sono responsabili della maggior parte delle funzioni della membrana. La maggior parte delle membrane sono composte dal 40% al 50% di lipidi (riferiti al peso), e dal 40% al 50% di proteine, la cui quantità e il cui tipo riflettono la funzione della membrana. Le membrane coinvolte nella trasduzione di energia, come le membrane interne dei mitocondri e le membrane tilacoidali dei cloroplasti, consistono fino al 75% di proteine. Alcune sono enzimi che catalizzano le reazioni associate alla membrana stessa, mentre altre, sono trasportatori che presiedono al trasporto di molecole e ioni specifici verso l’interno o l’esterno della cellula o dell’organello. Altre ancora agiscono come recettori e trasduttori di segnali chimici. Quindi la membrana plasmatica risulta costituita da proteine e lipidi in parti circa uguali (in peso), ma c’è anche un buon 10% di carboidrati, legati alle proteine e ai lipidi sulla superficie esterna. Molti carboidrati di membrana sono presenti sotto forma di glicoproteine e una piccola parte di glicolipidi, e cioè di lipidi con carboidrati a catena corta attaccati a essi. LIPIDI: I pricipali lipidi del plasmalemma sono i fosfolipidi, gli steroli ed in minor misura (10, 20%) i glicolipidi. I FOSFOLIPIDI : sono molecole anfipatiche, cioè costituite da una testa polare idrofila e da code apolari idrofobe. Questi si dispongono in modo da formare un doppio strato lipidico. La geometria risultante è controllata dalla forma delle molecole anfipatiche e dal bilancio dei domini idrofobici ed idrofilici. Quando messe a contatto con l’acqua queste molecole si autoassemblano spontaneamente in strutture di ordine superiore. Le teste idrofile massimizzano le loro interazioni con le molecole d’acqua, mentre le code idrofobiche interagiscono le une con le altre minimizzando la loro esposizione con la fase acquosa. I FOSFOLIPIDI : contengono una testa polare carica e due code idrocarburiche. Le code di acidi grassi contengono dai 14 ai 24 atomi di carbonio ed almeno una delle code ha uno o più doppi legami. Le pieghe introdotte dai doppi legami influenzano il loro impacchettamento che, a sua volta, influenza la fluidità complessiva della membrana. Steroli Fra gli steroli è comune e diffuso il ß-sitosterolo, mentre è più scarso il colesterolo(tipico componente delle membrane animali). Nel doppio strato lipidico gli steroli si comportano diversamente dai fosfolipidi, soprattutto perché il dominio idrofobico di una molecola di sterolo è molto più grande rispetto alla testa polare non carica. Di conseguenza gli steroli di membrana non solo sono capaci di diffondere rapidamente nel doppio strato, ma anche fare un “flip-flop” e con una frequenza superiore rispetto ai fosfolipidi. Molecola di colesterolo Il colesterolo è un fondamentale costituente della membrana plasmatica con l’importante ruolo di mantenerne la fluidità, soprattutto in presenza di agenti tendenti a fluidificarla (es. alcoli). Molecole di colesterolo all’interno del bilayer fosfolipidico Steroli presenti nella membrana I glicolipidi Pur non essendo marcatamente presenti, rivestono il ruolo di recettori, per mezzo della loro estremità glucidica sporgente rispetto alla membrana. I vari tipi di lipidi e fosfolipidi sono generalmente distribuiti in percentuali differenti in ognuno dei due strati lipidici della membrana, che può contenere oltre 100 tipi differenti di costituenti proteici. PROTEINE: Nella membrana plasmatica delle piante si trova in media il 3% delle proteine cellulari totali. Le proteine si associano con il doppio strato in differenti modi, ciascuno dei quali riflette la diversa funzione sia strutturale che enzimatica che esse svolgono. Le proteine infatti possono essere immerse in una o nell’altra faccia del doppio strato lipidico (proteine periferiche) oppure attraversarlo completamente(proteine integrali). Integrali monopasso Ancorate ai lipidi multipasso Periferiche SONO STATE IDENTIFICATE DUE CONFIGURAZIONI NELLE PROTEINE TRANSMEMBRANA: Una relativamente semplice di forma allungata, consistente in un’ α elica inserita nell’interno idrofobo della membrana, con porzioni idrofile non regolari che si protendono su entrambi i lati. Un’ altra viene individuata nelle proteine globulari a struttura tridimensionale complessa tale da formare ripetuti “passaggi”attraverso la membrana. Anche in queste proteine la catena polipetidica usualmente attraversa il doppio strato lipidico come una serie di α eliche. La proteina rappresentata a lato, possiede sette eliche transmembrana, (rappresentate schematicamente dai cilindri). I segmenti idrofili non elicoidali della proteina sono in contatto con le soluzioni acquose su entrambi i lati della membrana. Le zone lipofile interagiscono con le code lipofile delle molecole lipidiche,mentre le zone idrofile sono in connessione con le teste idrofile dei lipidi costituenti la membrana. Questa organizzazione rende stabile la struttura di membrana. Le proteine possiedono poli idrofili, nei quali si accumulano i gruppi polari dei componenti aminoacidici e poli lipofili formati da catene apolari di idrocarburi degli aminoacidi. Le membrane plasmatiche vegetali contengono un notevole numero di Vi sono anche dei flavocitocromi (NADH ossidasi e NADPH proteine con funzioni ossidoriduttive. ossidasi) transmembrana, presenti a bassi livelli e coinvolti Abbondanti enzimi redox del plasmalemma sono il citocromo b561 (che nelle reazioni di difesa da patogeni e nella riduzione dei avrebbe funzione di mantenere le comunicazioni tra l’ambiente cellulare e metalli. l’ambiente esterno) e alcune flavoproteine interne, come una NADH-cit b5 reduttasi e una chinone reduttasi NADPH-dipendente. La membrana plasmatica contiene inoltre numerosi recettori, ormonali e non,e il complesso proteico della cellulosa sintasi . L’unico complesso enzimatico intrinseco del plasmalemma vegetale che contribuisca alla sintesi di parete. I complessi enzimatici di cellulosa sintasi formano”rosette”incluse nella membrana plasmatica. Ogni”rosetta” sintetizza cellulosa da un derivato del glucosio, l’UDP-glucosio (uridina difosfato glucosio). Le molecole di UDP-glucosio penetrano nella rosetta dalla parte citoplasmatica della membrana e una microfibrilla viene estrusa dalla faccia esterna della membrana. Le proteine sono più voluminose e si muovono più lentamente,molte a causa del loro legame con il citoscheletro, sembrano essere praticamente immobili. LE MEMBRANE SONO FLUIDE Le membrane biologiche non sono strati statici di molecole rigidamente bloccate in una posizione. Una membrana è stabilizzata fondamentalmente da interazioni idrofobe,che sono molto più deboli dei legami covalenti. Il movimento laterale dei fosfolipidi all’interno della membrana è rapido, mediamente 2μm al secondo. Fluidità del doppio strato lipidico E’ fondamentale per i processi di trasporto e per le attività enzimatiche. Dipende da composizione in lipidi (lunghezza delle catene di acidi grassi e presenza di doppi legami) e dalla T . Temperatura di transizione di fase (Tm): t a cui il doppio strato artificiale si gelifica (congela) se raffreddato o diventa di nuovo fluido (fonde) se riscaldato. La membrana deve essere al di sopra di Tm per funzionare correttamente. Atomi di C e doppi legami… La membrana si mantiene fluida fino ad una certa temperatura (che dipende dal tipo di lipide di cui è costituita), al di sotto della quale i lipidi si associano tra di loro in modo particolarmente serrato,causandone la solidificazione. La membrana si mantiene fluida anche a basse temperature solo se è ricca di fosfolipidi con code idrocarburiche insature (i doppi legami impediscono loro di associarsi strettamente). Le singole molecole lipidiche in un doppio strato non sono legate covalentemente le une dalle altre, per cui sono libere di muoversi. Gli studi sul movimento dei fosfolipidi nei doppi strati hanno rilevato che queste molecole possono diffondere lateralmente, ruotare, flettere le loro code, oscillare su e giù e diffondere trasversalmente. La rotazione delle singole molecole attorno all’asse longitudinale ed il loro oscillare su e giù sono eventi molto rapidi. Contemporaneamente a questi moti, vi è il costante flettersi delle code idrocarburiche. Dato che la flessione aumenta verso la fine delle code, il centro del doppio strato ha il più alto grado di fluidità. Si ritiene che le singole molecole saltellano entro posti liberi (buchi) che si formano in seguito al moto termico delle molecole lipidiche di ciascun monostrato Flip-flop: un evento raro E’ invece sostenuta la rotazione e la diffusione laterale. Il “flip-flop”, avviene raramente. Si tratta infatti di un movimento energeticamente non favorito. Esistono cmq delle proteine particolari:”le flippasi” che mediano i movimenti dei lipidi neosintetizzati sempre in un’unica direzione. Meccanismo d’azione di un traslocatore di fosfolipidi L’asimmetria della membrana: si stabilisce durante la biogenesi nel reticolo endoplasmatico e viene mantenuta grazie alla presenza di traslocatori di membrana e quindi grazie alle differenti proteine presenti sui lati citosolico e extra-cellulare. Il movimento spontaneo di un lipide da uno strato all’altro (flip-flop) è raro. I trasporti di membrana avvengono secondo diverse modalità Trasporti: - attivi - secondariamente attivi caratteristiche contro-gradiente dipendenti dal metabolismo saturanti selettivi contro-gradiente accoppiati a flussi ionici saturanti selettivi - passivi facilitati secondo-gradiente saturanti selettivi - passivi diffusivi secondo-gradiente non selettivi non saturanti esempi Na/K-ATPase, Ca-ATPase, H/K-ATPase cotrasporto Na/glucosio, cotrasporto H/peptidi, antiporto Na/Ca trasporto di glucosio, trasporto di amminoacidi ioni attraverso i canali diffusione di sostanze lipofile Il passaggio di sostanze disciolte attraverso la membrana viene chiamato DIFFUSIONE. Grazie alla diffusione i soluti si distribuiscono uniformemente nelle soluzioni. La diffusione è rappresentata da un movimento netto di un soluto da una regione ad elevata concentrazione verso una a concentrazione inferiore. La velocità di diffusione varia a seconda della sostanza coinvolta, ma si mantiene comunque fino a quando non si è raggiunto l’equilibrio. Movimento di molecole attraverso le membrane: la diffusione semplice Permeabilità dei doppi strati lipidici Le molecole idrofile attraversano la membrana per mezzo di proteine di Membrana: trasportatori o canali Gli ioni diffondono attraverso i canali ionici Il flusso diffusivo di uno ione dipende dal Gradiente di concentrazione Gradiente elettrico a cavallo della membrana Trasporto Attivo • È richiesta energia metabolica (ATP) • I soluti si muovono CONTRO un gradiente di potenziale elettrochimico Tipi di trasporto attivo Trasporto attivo primario Trasporto attivo secondario Necessità di Energia Non richiede ulteriore energia oltre a quella derivata dal movimento molecolare Richiede ATP ENDOCITOSI DIFFUSIONE ESOCITOSI Diffusione Semplice Diffusione Facilitata Molecole attraversano il doppio strato lipidico Trasporto Attivo SECONDARIO Crea gradienti per Trasporto mediato da proteine di membrana Caratteristiche Strutturali FAGOCITOSI Trasporto Attivo PRIMARIO Vescicole rivestite da membrana
Scaricare