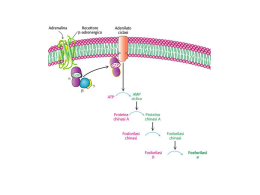
12/05/15 Le vie BIOSINTETICHE non sono mai s e m p l i c e m e n t e l ’ i nve r s o d e l l e corrispondenti vie CATABOLICHE I tessuti animali devono essere in grado di sintetizzare glucosio a partire da altri precursori per il funzionamento di tessuti ed organi che vivono solo di questa sostanza ed anche per l’immagazzinamento del glicogeno in altri tessuti. ORGANISMO CERVELLO 160 gr al dì 120 gr al dì RISERVE 190 gr nei fluidi corporei 20 gr Durante periodi di digiuno di durata superiore ad un giorno, il glucosio deve essere sintetizzato da altri precursori che non siano le riserve di glicogeno ormai terminate. Ciò è vero anche in una intensa attività fisica 1 12/05/15 FORMAZIONE DI GLUCOSIO DA PRECURSORI NON GLUCIDICI PRINCIPALI PRECURSORI SONO: lattato, aminoacidi, alanina prodotta dal muscolo attraverso il ciclo piruvato- alanina, propionato (degradazione di alcuni a.a) e il glicerolo L’organo in cui avviene la maggior parte della gluconeogenesi è il FEGATO sebbene in minor quantità avvenga anche nella midollare dei reni ma anche l’intestino tenue può essere una fonte di glucosio negli stati di digiuno. Avviene nel citosol anche se alcuni precursori sono prodotti nei mitocondri. Un difetto della gluconeogenesi è di solito fatale. L’ipoglicemia causa disfunzione cerebrale che può portare al coma e alla morte 2 12/05/15 Sette reazioni della GLICOLISI (reversibili) fanno parte anche della via GLUCONEOGENICA. Le tre reazioni irreversibili della glicolisi, catalizzate dall’ESOCHINASI, FOSFOFRUTTO CHINASI; PIRUVICO CHINASI, sono sostituite da diverse reazioni catalizzate da DIFFERENTI ENZIMI La gluconeogenesi utilizza enzimi specifici per aggirare tre reazioni irreversibili della glicolisi 3 12/05/15 PRIMA REAZIONE ESCLUSIVA 2 REAZIONI ESOERGONICHE 4 12/05/15 La piruvato carbossilasi REAZIONE ANAPLEROTICA catalizza la conversione ATP Utilizzata per mantenere ad un livello e Biotina dipendente del costante gli intermedi del ciclo di Krebs piruvato ad ossalacetato L’enzima richiede in modo assoluto l’acetil-CoA come attivatore allosterico La fosfoenolpiruvato carbossichinasi catalizza la reazione GTP dipendente dell’ossalacetato a fosfoenolpiruvato GTP al posto dell’ATP come donatore di energia nel fegato e nel rene dove si produce GTP nel ciclo di Krebs La reazione a fosfoenolpiruvato avviene nel citosol cellulare. Per essere trasformato in PEP nella seconda reazione l’ossalacetato deve attraversare la membrana mitocondriale che è impermeabile all’ossalacetato ma non lo è al malato CO2 fissata dalla piruvico carbossilasi è rilasciata dalla fosfoenolpiruvato carbossichinasi La formazione di ossalacetato nel citosol ha la funzione di fornire NADH necessario in altri passaggi della gluconeogenesi 5 12/05/15 SECONDA REAZIONE ESCLUSIVA Fruttosio-1,6-bisfosfatasi Fruttosio-1,6-bisfosfatasi Fruttosio-1,6-bisfosfato + H2O fruttosio-6-fosfato + Pi KJ/mole ΔG°’= -16,3 6 12/05/15 TERZA REAZIONE ESCLUSIVA Glucoso 6-fosfatasi Glucoso-6fosfatasi enzima localizzato principalmente nel reticolo endoplasmatico degli epatociti e nelle cellule renali 7 12/05/15 AMINOACIDI GLUCOGENICI 8 12/05/15 PROPIONATO a. Grassi o dall’ossidazione di alcuni a.a. GLICEROLO La regolazione della gluconeogenesi è cruciale per molte funzioni fisiologiche, ma in particolare per un corretto funzionamento del tessuto nervoso. Il controllo è particolarmente importante quando un animale si adatta all’esercizio muscolare o quando va incontro ad alternanza di condizioni di abbondante alimentazione o di digiuno G l u c o n e o g e n e s i e g l i c o l i s i r e a z i o n i i nve r s e avvengono in gran parte nel citosol e devono perciò essere regolate in modo reciproco. La regolazione reciproca è in gran parte legata alla carica energetica degli adenilati. 9 12/05/15 Anche l’acetil-CoA può essere visto come un regolatore reciproco della glicolisi e della gluconeogenesi, in quanto agisce sugli enzimi che interconvertono il piruvato e il fosfoenolpiruvato attivatore + inibitore La glucosio-6-fosfatasi non è regolata allostericamente , ma la sua Km per il glucosio-6fosfato è molto più alta della concentrazione intracellulare di questo metabolita per cui la v di reazione è di ordine uno Enzima di membrana del reticolo endoplasmatico 10 12/05/15 Il glucagone, ormone che segnala i bassi aa livelli di glucosio nel sangue provoca una diminuzione dei livelli di fruttosio 2,6bisfosfato aa rallentando nel fegato il consumo di glucosio da parte della glicolisi e stimolando la produzione di glucosio per l’espor tazione da par te della gluconeogenesi. Nel fegato la regolazione reciproca della fosfofruttochinasi e della fruttosobisfosfatasi impedisce un ciclo futile e regola la concentrazione di glucosio nel sangue La carica energetica influisce sulla regolazione della glicolisi e della gluconeogenesi agendo sulla interconversione del fruttosio-6-fosfato e fruttosio-1,6-bisfosfato e determinando un aumento o una diminuzione di ATP a seconda delle esigenze della cellula . L’AMP stimola le vie che ossidano i nutrienti e inibisce la gluconeogenesi, per fornire ENERGIA alla cellula epatica 11 12/05/15 Effettore allosterico i cui livelli riflettono i livelli di glucagone nel sangue e quindi regolatore del glucosio ematico Regolazione dei livelli EMATICI di glucosio Il fruttosio 2,6-bisfosfato è una molecola di fruttosio che viene prodotta e defosforilata in posizione 2 da uno stesso complesso proteico che contiene due attività enzimatiche che catalizzano reazioni inverse PFK-2 FBPasi-2 12 12/05/15 La concentrazione di fruttosio-2,6-bisfosfato dipende dalla velocità della sua formazione e distruzione. Quando il glucagone arriva sulla membrana di un epatocita determina la formazione di cAMP che attiva una proteina chinasi a catalizzare la fosforilazione del complesso proteico determinando l’attivazione enzimatica della FBPasi-2 e l’inattivazione contemporanea della PFK-2. Attività distinte di una stessa proteina PFK-1 FBPasi-1 La fosforilazione da parte della proteina chinasi diminuisce l‘attività della PFK-2 ed aumenta quella della FBPasi-2 La formazione del fruttosio-2,6-bis-P è regolata covalentemente attraverso una cascata enzimatica innescata dal Glucagone 13 12/05/15 INOLTRE controlla i livelli dell’enzima FOSFOENOLPIRUVATO CHINASI attivandone la trascrizione del gene strutturale; mentre deprime la sintesi della PIRUVATO CHINASI INSULINA effetto opposto Il livello di glucosio nel sangue aumenta grazie a due meccanismi: 1. Regolazione ad opera del cAMP della regolazione a cascata che porta alla demolizione del glicogeno. 2. La depressione sempre ad opera del cAMP, del livello del fruttosio-2,6-bisfosfato, che stimola la gluconeogenesi 14 12/05/15 Un compito importante nella regolazione del glucosio nel sangue è dovuto, oltre ovviamente all’attività dei diversi ormoni, alla capacità dei trasportatori di glucosio e delle loro differenze nelle diverse cellule. Punti d’ingresso degli amminoacidi Altri substrati per la gluconeogenesi Effetti allosterici Importante nei ruminanti 15 12/05/15 16 12/05/15 1. formazione metabolicamente irreversibile Il substrato per la sintesi del glicogeno catalizzata dalla GLICOGENO SINTASI è l’UDP-glucosio SINTESI DELL’UDP-glucosio Molte delle reazioni in cui gli esosi vengono trasformati o polimerizzati utilizzano 2. Aumentano l’efficienza catalitica dell’enzima 3. I gruppi nucleotidilici sono eccellenti gruppi uscenti 4. “etichetta” molecolare 1. ZUCCHERI LEGATI A NUCLEOTIDI SINTESI DEL GLICOGENO Avviene praticamente in tutti i tessuti animali, ma soprattutto nel fegato e nel muscolo scheletrico Glucosio-6fosfatasi Una parte di glucosio ingerito durante il pasto segue una via più lunga: entra negli eritrociti dove viene convertito in lattato dalla glicolisi, esce quindi nel torrente sanguigno come lattato va al fegato dove sarà convertito in glucosio-6fosfato nella gluconeogenesi per la sintesi di glicogeno. 17 12/05/15 Se la biosintesi del glicogeno dipendesse unicamente dalla glicogeno sintasi il glicogeno sarebbe costituito da lunghe catene lineari simili a quelle dell'amilosio. La GLICOGENO SINTASI catalizza il trasferimento di residui UDP-glucoso alle estremità non riducenti di una ramificazione del GLICOGENO costituita da almeno 8 residui di glucosio Formazione delle ramificazioni ENZIMA RAMIFICANTE denominato anche Amilo (1,4->1,6) transglicosilasi in quanto rompe un legame α1,4 e forma un legame α-1,6 Tale enzima consente lo spostamento di un oligomero di 6-8 residui di glucosio (legati con legame α- 1,4) dall'estremità non riducente di una catena ad un residuo di glucosio , lontano almeno 4 residui dalla ramificazione più vicina Le ramificazioni aumentano la solubilità del polimero e aumenta, inoltre, il numero delle estremità non riducenti che sono i siti per la demolizione e la sintesi del glicogeno 18 12/05/15 Ma la glicogeno fosforilasi non è capace di legare due molecole di UDP-glucosio per dare inizio alla sintesi del glicogeno Il primer per la Glicogeno sintasi è rappresentato da una corta catena di residui di glucosio sintetizzata da una proteina chiamata GLICOGENINA, che trasferisce il glucosio dall’UDP-Glc ad un residuo di tirosina della stessa proteina. Inneschi di circa 8 residui che vengono in seguito estesi dalla Glicogeno sintasi. La glicogenina continuerà a far parte della molecola di glicogeno restando legata covalentemente all’estremità a cui ha dato origine 19 12/05/15 Le condizioni che attivano la demolizione del glicogeno inibiscono la sintesi del glicogeno e viceversa L’adrenalina e il glucagone inibiscono la sintesi del glicogeno con gli stessi meccanismi con cui ne promuovono nello stesso tempo demolizione 20 12/05/15 Proteina chinasi A cAMP dipendente (attiva) 21 12/05/15 22
Scaricare