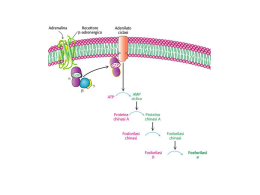
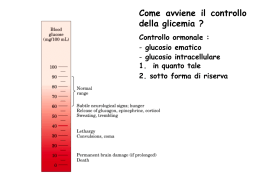
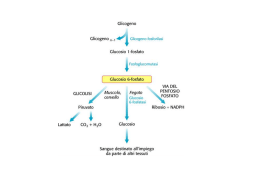
Biologia Molecolare della Cellula ’08 7: Regolazione ormonale del metabolismo del glicogeno e della glicolisi Desensitizzazione cAMP Proteina-CHINASI cAMP DIPENDENTE (PKA) Proteine target : ε metabolici Fosforilasi chinasi Glicogeno Fosforilasi Glicogeno Sintasi Diminuzione della sintesi del glicogeno Aumento della demolizione del glicogeno I principali ormoni che regolano il metabolismo energetico negli animali Ormone Insulina Glucagone Epinefrina Effetti biochimici Permeabilita’ cellulare (muscolo e tessuto adiposo) Glicolisi Sintesi del glicogeno Sintesi dei trigliceridi Gluconeogenesi Lipolisi Degradazione delle proteine Sintesi proteica, del DNA e RNA Livelli di cAMP (fegato e tessuto adiposo) Glicogenolisi Sintesi del glicogeno Idrolisi dei trigliceridi Gluconeogenesi Glicolisi Livelli di cAMP nel muscolo Mobilizzazione dei trigliceridi Glicogenolisi Sintesi del glicogeno Effetti fisiologici Segnala lo stato di abbondanza di nutrienti Livello di glucosio ematico Immagazzinamento dei combustibili Crescita cellulare e differenziamento Rilascio di glucosio dal fegato Livello di glucosio ematico Rilascio di glucosio dal fegato Utilizzo del glucosio da parte del muscolo Livello di glucosio ematico Gli ormoni chiave nella regolazione del metabolismo energetico sono l’insulina, che promuove l’utilizzo del glucosio, e il glucagone e l’epinefrina, che aumentano il livello ematico del glucosio CONTROLLO del GLUCOSIO EMATICO (glicemia) β-cells α-cells Negli animali il glucosio viene immagazzinato sotto forma di GLICOGENO (un grosso polimero del glucosio) nel fegato e nel muscolo scheletrico. Nel fegato il glucosio ottenuto dalla degradazione del glicogeno puo’ essere rilasciato nella circolazione perche’ possa essere assorbito dagli altri tessuti. Nel muscolo viene utilizzato per generare energia necessaria per la contrazione. 6 4 1 6 6 5 5 1 1 4 4 3 2 3 2 Legame α-1,4 glucosidico glicogeno CONTROLLO del GLUCOSIO EMATICO (glicemia) NELSON cap. 15 Funzione del fegato e’ di mantenere costante (circa 5mM) la concentrazione di glucosio nel sangue. Risponde a GLUCAGONE e insulina. - bassa [gluc] (= fame, attivita’) α cells pancreas rilasciano GLUCAGONE aum. cAMP demolizione del glicogeno e inibiz. della glicolisi - alta [gluc] β cells pancreas rilasciano INSULINA attivazione PP1 SINTESI del GLICOGENO La PKA nel METABOLISMO del GLICOGENO SINTESI del glicogeno: UDP-glucosio + glicogeno (n residui) DEGRADAZIONE: glicogeno (n) glicogeno FOSFORILASI glicogeno SINTASI glucosio1- glicogeno (n+1) + UDP + glicogeno (n-1) Pi muscolo e altri tessuti il glucosio 6-P entra nella glicolisi MUTASI gluc1- gluc 6- fegato : gluc1- MUTASI gluc 6- glicolisi G-6-FOSFATASI glucosio ATP SANGUE altri tessuti Pi L’aumento della concentrazione di cAMP indotta da stimolazione con adrenalina o glucagone aumenta la conversione da glicogeno a glucosio1- mediante: - L’inibizione della SINTESI del glicogeno - L’attivazione della sua DEGRADAZIONE (glicogenolisi) Il processo e’ reversibile quando cala la concentrazione di cAMP a Sintesi e degradazione del glicogeno Legame α-1,4 glucosidico b GLICOGENO FOSFORILASI (GP) GLICOGENO SINTASI (GS) (a) L’Incorporazione del glucosio da UDP-glucosio nel glicogeno e’ catalizzato dalla glicogeno sintasi. (b) La rimozione di unita’ di glucosio dal glicogeno e’ catalizzata dalla glicogeno fosforilasi. Siccome due enzimi differenti catalizzano la formazione e la degradazione del glicogeno, le due reazioni possono essere regolate in modo indipendente. (R e’ la porzione rimanente della molecola di glicogeno) LODISH cap.20 Regolazione della sintesi e degradazione del glicogeno indotta da cAMP in cellule epatiche e muscolari GLUCAGONE PKA Ca2+ PIP- (a) Un aumento del cAMP citosolico attiva la chinasi cAMP dipendente (cAPK-PKA) che innesca una cascata di protein chinasi che include la Glicogeno fosforilasi chinasi (GPK) e la Glicogeno fosforilasi (GP), e porta alla demolizione del glicogeno. La cAPK attiva fosforila e inattiva la Glicogeno Sinatasi (GS) inibendo la sintesi del glicogeno. La fosforilazione di un inibitore (PIP) della fosfoprotein fosfatasi (PP) (v. oltre) impedisce alla PP di defosforilare gli enzimi attivati della cascata di chinasi e la GS inattiva. (Gli enzimi attivi sono in toni di colore piu’ scuri, i toni piu’ chiari corrispondono alla forma inattiva) Regolazione della sintesi e degradazione del glicogeno indotta da cAMP in cellule epatiche e muscolari (INSULINA) Fosfatasi (b) Un decremento del cAMP inattiva la PKA, e libera la fosfoprotein fosfatasi (PP) nella sua forma attiva. Questo enzima rimuove i fosfati dalla GPK e GP, inibendo quindi la demolizione del glicogeno. La fosfatasi rimuove anche il fosfato alla glicogeno sintasi (GS) inattiva, attivando questo enzima e stimolando la sintesi del glicogeno. (Gli enzimi attivi sono in toni di colore piu’ scuri, i toni piu’ chiari corrispondono alla forma inattiva) Le Protein fosfatasi Phosphoprotein Phosphatase (PP) Protein fosfatasi 1 (PP-1) Protein fosfatasi 2A (PP2A) Protein fosfatasi 2B Protein fosfatasi 2C PP-1 ATTIVA cAMP PP-1 INATTIVA PKA PIP attivo P PIP attivo P ADP ATP PIP inattivo Glicogeno fosforilasi (GP) Fosforilasi chinasi (GPK) Glicogeno sintasi (GS) La protein fosfatasi-1 e’ inibita da un inibitore fosforilato: PIP (Phosphatase Inhibitor Protein) L’aumento del cAMP inibisce l’attivita’ della protein fosfatasi 1 (PP-1) in quanto la PKA rende attivo l’inibitore, fosforilandolo La fosforilazione da parte della PKA di - Glicogeno FOSFORILASI CHINASI (GPK) (attivattore glicogeno fosforilasi) - Glicogeno SINTASI ha effetti opposti sull’attivita’ dei due enzimi assieme all’inibizione della fosfatasi porta alla mobilitazione del glicogeno GPK GPK- GS INATTIVA ATTIVA GP GP- GS- SINTESI DEGRADAZIONE gluc 1P Quando cala la concentrazione di cAMP prevale l’attivita’ della Protein fosfatasi (PP) con effetto opposto HR cAMP UDP UDP- Gluc PKA R2C2 R2 2C GPK DEGRADAZIONE glicogeno GS GS 4cAMP SINTESI GP Gluc-1P GP Ca2+ GPK HR nel fegato = GLUCAGONE (ADRENALINA) nel muscolo = ADRENALINA attivazione inibizione ( => GPCR accoppiati a Gs) CONTROLLO della GLICOLISI da PKA Nel muscolo il G1P prodotto dalla GP e’ utilizzato per generare ATP G1P G6P glicolisi ciclo acido citrico Nel fegato il glucosio puo’ essere rilasciato nel circolo sanguigno in quanto e’ presente glucosio-6-fosfatasi, G1P G6P G6 fosfatasi glucosio + Pi e grazie al blocco della glicolisi. La regolazione della glicolisi avviene a livello delle 3 reazioni esoergoniche, mediante regolazione allosterica e modificazioni covalenti. La Fosfofruttochinasi 1 (PFK-1) e’ un enzima chiave F6P + ATP F1,6BP + ADP Tetramero di 340KDa Inibito da ATP, CITRATO Inibizione da ATP e’ reversibile da AMP Inoltre e’ attivata da F2,6BP La regolazione mediata da cAMP dell’attivita’ PFK-1 avviene a livello della concentrazione di F2,6BP GLICOGENO GP Gluc.1P PFK-1 mutasi Regolazione ormonale della glicolisi nel fegato glicogeno sangue GP GLUCOSIO GS UDP UTP Il fruttosio 2,6 bisfosfato (F2,6BP) e’ regolatore della glicolisi. Riflette la disponibilita’ di GLUCOSIO = Basso GLUCAGONE [F2,6BP] Glicolisi [F2,6BP] Glicolisi G1P G6P F6P PFK-2 F2,6BP FBPase-2 PFK-1 FBPase-1 F1,6BP PEP Gluconeogenesi PIRUV attivazione inibizione SINTESI e DEGRADAZIONE del F2,6BP sono REGOLATE da FOSFORILAZIONE dell’ENZIMA TANDEM (PFK-2 / FBPase-2) PFK -2 FBPase -2 enzima tandem Fosfofrutto chinasi-2/Fruttosio Bisfosfatasi- 2 PKA Fosforilazione PFK2/FBPase2 (= glucagone, BASSO glucosio) ATTIVAZIONE FBPase-2 INIBIZIONE PFK-2 F2,6BP Controllo del metabolismo del GLUCOSIO da cAMP in fegato Glicogenolisi Glicolisi La modulazione della trasmissione del segnale a livello del RECETTORE: la DESENSITIZZAZIONE - Spegnimento del segnale -Adattamento alla intensita’ dello stimolo Permette di ottenere una risposta cellulare che dipende dalla variazione dell’intensita’ dello stimolo piuttosto che alla quantita’ assoluta dello stesso risposta correlata alla Δ[H] quando la stimolazione e’ prolungata Riduzione del numero di recettori funzionali per cellula effetto a breve o a lungo termine (es. inattivazione funzionale / sequestro del recettore oppure degradazione) per stimoli ormonali, neurotrasmettitori o stimoli sensoriali DESENSITIZZAZONE La quantita’ di una specie di recettore sulla superficie cellulare non e’ costante: il suo livello e’ modulato - SENSITIZZAZIONE (incremento del n.recettori) - DESENSITIZZAZIONE (riduzione del n. recettori funzionali) L’esposizione prolungata all’ormone porta alla riduzione dei recettori funzionali. Cio’ DESENSITIZZA la cellula a quel dato ligando = per ottenere la medesima risposta fisiologica e’ necessario uno stimolo (conc. ligando) piu’ elevato. • I recettori possono essere internalizzati per endocitosi e degradati • I recettori possono essere internalizzati, ma restano in vescicole intracellulari e possono essere riciclati in superficie (sequestro) • I recettori restano in superficie ma diminuisce la loro affinita’ per il ligando • I recettori restano in superficie, legano l’ormone, ma il complesso ormone recettore non e’ piu’ funzionale - disaccoppiamento dal sistema di trasduzione del segnale (es. non e’ piu’ in grado di riconoscere / attivare le proteine G) SEQUESTRO DOWN-REGOLAZ INATTIVAZIONE RECETTORE INATT PROTEINA SEGNALATRICE PRODUZIONE INIBITORE Cinque modi in cui una cellula target puo’ divenire desensitizzata a una molecola segnale. I meccanismi di inattivazione che sono mostrati sia per il recettore che per le proteine segnalatrici avvengono spesso mediante fosforilazione della proteina che viene inattivata, ma ne sono conosciuti anche altri. Nella chemiotassi dei batteri la desensitizzazione dipende dalla metilazione del recettore. Il processo reversibile di ADATTAMENTO o DESENSITIZZAZIONE permette alle cellule di rispondere a cambiamenti della concentrazione di ligando, piuttosto che alla concentrazione assoluta dello stesso, per una finestra piuttosto ampia di concentrazione di ligando. - uno o piu’ meccanismi possono essere coinvolti - spesso coinvolge la fosforilazione dei recettori - un effetto secondario e’ l’abbassamento della [H] libero nell’ambiente extracellulare (per legame non produttivo) Rec. β2-Adrenergico Siti di GLICOSILAZIONE (S/T X N) SX N C C INTERAZIONE con il LIGANDO D113 sulla TM3 S204, S207 sulla TM5 F290 sulla TM6 D RRX S PONTI DISOLFURO S Siti di FOSFORILAZIONE PKA (R K/R X S/T) Siti di FOSFORILAZIONE βARK (beta Adrenergic Receptor Kinase) L’Attivazione della PKA downregola i recettori accoppiati a Gs Diagramma schematico della regolazione a feedback che controlla l’attivita’ di recettori accoppiati a Gs mediante cicli di fosforilazione e defosforilazione. Tutti i recettori di questo tipo sono fosforilati dalla chinasi cAMP dipendente (PKA). Residui addizionali sono fosforilati da chinasi specifiche per il recettore, come βARK, il cui substrato e’ il recettore beta-adrenergico, o da GRK. Chinasi ATTIVATA dal RECETTORE BARK (βARK ) = Beta Adrenergic Receptor Kinase Gβγ Kinase PH Domini di Interazione I domini PH (Pleckstrin Homology) N Costituiti da circa 100 aa 2 β-sheet antiparalleli β-strands connessi da loop non conservati α-elica C-terminale C Funzione e legame del dominio: • I domini Pleckstrin-homology (PH) si ritrovano in molte proteine della trasduzione del segnale che si associano alle membrane. • Alcuni domini PH legano con alta affinita’ fosfoinositidi specifici formati durante la trasduzione del segnale (PI-4,5P2; PI-3,4P2, PI-3,4,5P3 - Kd a bassa µM, o nM). • Il legame ai fosfoinositidi potrebbe permettere alle proteine con domini PH di rispondere ai messaggeri lipidici per esempio mediante la rilocalizzazione presso la membrana. • La porzione C-ter di alcuni PH e’ in grado di legare le subunita’ beta/gamma di G proteine eterotrimeriche (v. BARK).
Scaricare