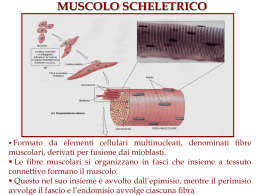
TESSUTO MUSCOLARE SCHELETRICO TESSUTO MUSCOLARE Il tessuto muscolare è caratterizzato dalla proprietà contrattile particolarmente sviluppata legata alla presenza di filamenti contrattili: actina e miosina Esistono 3 tipi di tessuto muscolare: Tessuto muscolare striato scheletrico (volontario) Tessuto muscolare striato cardiaco (involontario) Tessuto muscolare liscio (involontario) • le dimensioni della fibra muscolare possono raggiungere anche qualche cm di lunghezza, mentre il diametro può variare da qualche decina a qualche centinaio di µm • la fibra muscolare presenta: – il sarcolemma – all'esterno c'è un ulteriore rivestimento di glicoproteine che risulta PAS positivo, ma non è compreso nella definizione di sarcolemma – i nuclei (fino a una quarantina per mm di lunghezza) disposti alla periferia (subsarcolemmali) – il sarcoplasma che contiene a sua volta: • le miofibrille (elementi bastoncellari che hanno lunghezza pari alla lunghezza della fibra e diametro di 1µm ; sono costituiti da fascetti di filamenti) • i mitocondri • il reticolo liscio sarcoplasmatico) (è specializzato e si dice reticolo Istogenesi Il TMSS non è costituito da singole cellule ma da cellule fuse tra di loro, i mioblasti. I mioblasti originano dal miotomo e migrano per costituire i muscoli del tronco e degli arti. I mioblasti proliferano ulteriormente, si dispongono in serie e fondono le proprie membrane formando il miotubo, rivestito da un unica membrana e contenente il citoplasma e i nuclei dei mioblasti (meccanismo sinciziale). Alcuni mioblasti rimangono all’esterno della membrana del miotubo e costituiscono una riserva per necessità rigenerative o riparative del muscolo (cellule satelliti). All’interno del miotubo si ha un’intensa sintesi di proteine miotomo contrattili che si organizzano in miofilamenti i quali a loro volta si miotubo aggregano in miofibrille queste ultime giungono a diventare così abbondanti da sospingere i nuclei verso la periferia dell’ elemento muscolare Si è costituita la fibra muscolare, entità morfologica costitutiva del muscolo scheletrico rivestita da un'unica membrana (sarcolemma) un qualsiasi muscolo dell’organismo è una struttura formata da parecchie fibre muscolari le fibre muscolari vengono tenute assieme dal tessuto connettivo la capsula di connettivo fibrillare denso che avvolge il tessuto muscolare prende il nome di epimisio dall’epimisio si dipartono setti connettivali che scompongono il muscolo in tanti fascetti di fibre muscolari questo connettivo diventa sempre più delicato (connettivo lasso) e prende il nome di perimisio; al suo interno si ritrovano vasi arteriosi, venosi, linfatici e nervi le parti ancora più interne del perimisio si continuano con una delicata rete di connettivo ancora più sottile di tipo reticolare che forma un rivestimento intorno ad ogni singola fibra muscolare quest’ultimo rivestimento prende il nome di endomisio epimisio perimisio endomisio 1 TESSUTO MUSCOLARE SCHELETRICO Morfologia della fibra muscolare al M.O. in sez. trasversale Morfologia della fibra muscolare al M.O. in sez. longitudinale • in sezione trasversale forma poligonale • Ø pressoché costante • Caratteristiche della fibra muscolare in sezione trasversale: – Presenti più nuclei e in posizione periferica – l’interno della fibra appare punteggiato per la sezione delle miofibrille • Striatura trasversale per alternanza di dischi chiari e dischi scuri • Presenti più nuclei schiacciati e in posizione subsarcolemmale miofibrilla Fibra muscolare La striatura appare omogenea perché le miofibrille sono: 1 numerose; 2 fittamente stipate fra di loro; 3 disposte fra loro in fase o in registro. Il disco scuro si definisce DISCO A (anisotropo) Il disco chiaro si definisce DISCO I (isotropo) Questa definizione è per il comportamento dei dischi nei confronti della luce polarizzata A forte ingrandimento si osservano: Banda A Banda I Linea z a metà del disco chiaro una sottile linea detta linea Z Nella parte centrale del disco scuro una zona con densità intermedia tra quella del disco chiaro e quella delle parti laterali del disco scuro. Questa banda è la banda H. Nella parte centrale del disco scuro nella sua posizione mediana si trova la linea M Banda I Banda A Linea Z Il disco anisotropo provoca deviazione del fascio di luce polarizzata e appare più luminoso Ciò dimostra che al suo interno c’è una maggiore presenza di strutture proteiche fortemente organizzate Il disco I risulta scuro poiché non devia il fascio di luce polarizzata 2 TESSUTO MUSCOLARE SCHELETRICO IL SARCOMERO Ciascuna miofibrilla è data dal ripetersi di segmenti uguali fra loro. Questo segmento è il sarcomero sarcomero, unità morfo morfo-funzionale della miofibrilla. È compreso fra due linee Z successive Ha una lunghezza di 2,5 – 3µm a riposo È costituito da un emidisco chiaro, un disco scuro, un emidisco chiaro ULTRASTRUTTURA filamenti di actina o miofilamenti sottili • Ø di 5-6nm • lunghezza di 1µm • costituiti essenzialmente da actina filamenti di miosina o miofilamenti spessi • Ø di 15nm • lunghezza di circa 1,5µm • costituiti essenzialmente da miosina La disposizione dei due tipi di filamenti è responsabile della diverse densità che si trovano all’interno del sarcomero I filamenti sottili di actina si ancorano da ciascun lato sulle linee Z e da qui convergono verso il centro del sarcomero senza raggiungerlo in quanto si arrestano al confine tra parte laterale del disco scuro e banda H I filamenti spessi di miosina occupano tutta la lunghezza del disco scuro i dischi chiari chiari, contengono solo i filamenti sottili, saranno necessariamente la parte meno densa di tutto il sarcomero le parti laterali dei dischi scuri scuri, contengono entrambi i tipi di filamenti, saranno la parte più densa di tutto il sarcomero la banda H contiene solo i filamenti spessi, avrà una densità minore rispetto a quella delle parti laterali del disco scuro, ma maggiore di quella dei dischi chiari sezione trasversale a livello dell’emidisco I mostra mostra:: punti del diametro di 6nm corrispondenti ai filamenti sottili; disposti ai vertici di ipotetici esagoni regolari sezione trasversale a livello della banda H punti più grandi dei precedenti, del diametro di 15nm corrispondenti ai filamenti di miosina; disposti ai vertici di ipotetici triangoli equilateri sezione trasversale a livello delle parti laterali del disco scuro Sono presenti entrambi i tipi di filamenti che mantengono la disposizione spaziale contrazione Ciascun filamento spesso si trova al centro di un esagono e ciascun filamento sottile al centro di un triangolo equilatero Questa disposizione così rigorosa risponde all’esigenza di permettere ai due tipi di filamenti di interagire tra di loro per garantire il meccanismo della contrazione 3 TESSUTO MUSCOLARE SCHELETRICO I filamenti di actina i filamenti sottili o di actina di spessore di circa 6nm e lunghezza di 0,8-1µm, si dipartono da ciascuna linea Z e si portano verso il centro del sarcomero terminando al confine tra parte laterale del disco scuro e banda H composizione molecolare ciascun filamento è costituito da diverse proteine ma la sua struttura molecolare principale è rappresentata dalla actina questa non differisce grandemente dall’actina dei microfilamenti citoscheletrici non muscolari (è una isoforma detta actina sarcomerica) esiste in forma globulare o G-actina che polimerizza e costituisce delle strutture filamentose (F-actina) molecole di F-actina si uniscono a due a due cosicché il filamento sottile risulta dall’assemblaggio di due F-actina avvolte tra di loro a spirale altre proteine del filamento sottile in rapporto con la linea Z Proteine regolative del filamento sottile Nel filamento sottile sono presenti altre proteine definite accessorie o regolative: la tropomiosina B e la troponina tropomiosina B proteina filamentosa, nastriforme la cui molecola raggiunge una lunghezza di circa 40nm Le molecole di tropomiosina si inseriscono, in successione, nel passo della spirale formata dalle F-actine troponina proteina globulare a forma di ferro di cavallo che si dispone a cavallo del filamento sottile a distanze regolari di 40nm la troponina è scomponibile in 3 subunità la troponina T: T per tropomiosina perché è la subunità che può legarsi alla tropomiosina la troponina I dove I sta per inibitoria capace di legare la G actina Troponina C, è interposta tra le altre due , dove C sta per calcio a cui si lega I filamenti spessi nebulina: si stratifica sul filamento impedendo depolimerizzazione e polimerizzazione il filamento spesso o di miosina, Ø di circa 15nm, lunghezza di circa 1,5µm, occupa tutta l’estensione del disco scuro da ciascun filamento sottile si originano 4 subfilamenti detti subfilamenti Z, diretti lungo gli spigoli di una piramide a base quadrangolare il cui vertice sta nell’origine del filamento sottile; ciascun subfilamento si connette con 1 subfilamento del sarcomero adiacente non necessariamente i 4 subfilamenti di un filamento di actina prendono rapporto con 4 subfilamenti di uno stesso filamento di actina del sarcomero adiacente esiste una disposizione più complessa responsabile dell’andamento a zig zag della linea Z nelle parti laterali del disco scuro presenta delle appendici laterali, dette ponti trasversi, che sporgono dalla superficie del filamento stesso di circa 7nm, trasversi angolate di circa 90° rispetto all’ asse maggiore del filamento la parte centrale del filamento stesso, per un’ ampiezza di circa 250nm è priva di questi ponti trasversi la molecola principale del filamento è la miosina alla diffrattografia ai raggi X la lunghezza è di 150nm e uno spessore di 2-7nm. Ponte trasverso Ponte trasverso I subfilamenti sono costituiti da alfa actinina che presenta siti di legame per la titina o connectina e da tropomiosina A che è un isomero della tropomiosina B 4 TESSUTO MUSCOLARE SCHELETRICO La molecola di miosina Presenta una parte bastoncellare (coda coda) Ø circa 2nm e una parte globosa (testa testa) con ampiezza di circa 7nm situata all’altra estremità (corrisponde ai ponti trasversi del filamento spesso) la molecola di miosina è assimilabile ad una mazza da golf Mediante digestione enzimatica con tripsina la miosina si scompone in due parti: meromiosina leggera (LMM) LMM) e questa corrisponde alla quasi totalità della parte bastoncellare meromiosina pesante (HMM) HMM): comprende un breve tratto della coda che fa seguito alla meromiosina leggera e l’intera testa globosa o ponte trasverso angolato di circa 90° e con un'estensione di 7nm contiene due catene pesanti e 4 catene leggere che risiedono solo nella testa globosa un’ulteriore digestione enzimatica con papaina permette di scomporre la meromiosina pesante in due subunità: S2: è la parte della meromiosina pesante che fa seguito alla meromiosina leggera S1: corrisponde alla testa globosa e contiene un sito per la actina globulare, ATP e ATPasi da ciascun lato del filamento spesso (in ciascuna metà del sarcomero) le molecole di miosina si dispongono con regolarità e ripetitività: ripetitività sono sfasate in senso lineare di 14, 14,3nm l’una rispetto alla successiva le teste sono disposte in modo elicoidale in un segmento di 42, 42,9nm sono compresi 6 ponti diretti verso i filamenti di actina disposti ai vertici dell’esagono che si trova attorno al filamento spesso Nel costituire il filamento spesso le molecole di miosina si dispongono con polarità opposta in prossimità della linea M Linea M Le prime molecole più vicine alla metà del sarcomero sia da un lato che dall’altro del filamento dispongono le code dirette verso la linea M e le teste globose dirette verso la linea Z. di uno o dell’altro lato Questa disposizione spiega perchè la parte centrale del filamento sia priva di ponti trasversi per un ampiezza di 250nm (somma della coda di una molecola di un lato con quella dell’ altro lato) La disposizione con polarità opposta è essenziale per il meccanismo della contrazione il filamento di miosina può stabilire rapporti con i filamenti di actina che gli stanno intorno per mezzo dei ponti trasversi delle molecole di miosina che lo compongono ZONE FLESSIBILI per stabilire tali rapporti si presuppone l’esistenza di zone flessibili tra S1 ed S2 e tra S2 e MML 5 TESSUTO MUSCOLARE SCHELETRICO Nel filamento di miosina in prossimità della linea M sono presenti altre proteine: Proteina M forma una rete in cui si inseriscono i filamenti spessi che così vengono mantenuti in posizione per mantenere le miofibrille in registro Miomesina e CPK (creatin-fosfochinasi muscolare) si trovano anch’esse in prossimità della linea M con funzioni di stabilizzare i filamenti. Possono legare altre proteine sarcomeriche (es. l’estremità M della titina) la titina: titina ha un peso molecolare che si aggira intorno ai 3.000 kD è una struttura molecolare elastica che si inserisce nel filamento spesso tra le molecole di miosina legandosi alla proteina C e ancorandosi alla linea M per mezzo della miomesina all’estremità del filamento di miosina rivolta verso la linea Z si continua al di fuori del filamento stesso e quindi viene tesa come un elastico tra filamenti spessi e linee Z che raggiunge passando tra i filamenti sottili cui si associa lateralmente si àncora alla linea Z legandosi all’a-actinina e alla telethonina Proteina C distribuita nella parte del filamento che presenta i Probabilmente lega la titina lungo il filamento spesso ponti trasversi. Proteina H presente nella banda H forse con funzione analoga alla proteina C il suo ruolo è quello di: stabilizzare la posizione dei filamenti nelle miofibrille impedire una sovradistensione del sarcomero durante il processo di rilasciamento ovvero di impedire alle linee Z di allontanarsi tra di loro oltre una certa misura CENTROTUBULI O TUBULI T E TRIADI DI PORTER E PALADE RETICOLO SARCOPLASMATICO Il reticolo sarcoplasmatico è fomato da una serie di tubuli più o meno ampi, i sarcotubuli I sarcotubuli costituiscono 3 formazioni: - le cisterne fenestrate - i tubuli longitudinali - le cisterne terminali Al confine fra disco A ed emidisco I i tubuli longitudinali confluiscono in una formazione che circonda ciascuna miofibrilla: la cisterna terminale A livello del confine tra banda A ed emisco I si trova un altra cisterna terminale dalla quale si originano altri tubuli longitudinali che decorrono parallelamente rispetto all’asse della miofibrilla passando sopra il disco chiaro Questi tubuli superano la linea Z e raggiungono il sarcomero successivo In questo caso i tubuli longitudinali dove, a livello di confine tra emidisco sono più corti e lungo il loro decorso I e disco A confluiscono in una nuova non c’è la cisterna fenestrata cisterna terminale Al confine tra disco chiaro e disco scuro c’è una coppia di cisterne terminali e tra le due cisterne si inserisce una struttura che deriva per invaginazione dal sarcolemma: il centrotubulo In prossimità del confine tra disco chiaro e scuro il sarcolemma si approfonda verso l’interno e forma dei tubicini che appena si trovano davanti ad una miofibrilla si dividono a 180° e per questo si chiamano anche tubuli T L’insieme dei centrotubuli costituisce il sistema T; le due diramazioni della T formano circonferenze che circondano ciascuna miofibrilla interponendosi tra le due cisterne terminali Non esiste una vera e propria continuità morfologica tra centrotubuli e cisterne terminali. L’insieme di due cisterne terminali e del centrotubulo compreso tra di loro costituisce la triade di Porter e Palade Ogni sarcomero è dotato di due triadi (ciascuna situata al confine disco chiaro-disco scuro) 6 TESSUTO MUSCOLARE SCHELETRICO LA SINAPSI NEUROMUSCOLARE O PLACCA MOTRICE La fibra completa si porta verso la periferia dell’organismo e raggiunge il muscolo dove deve portare il segnale per la contrazione. In prossimità del muscolo la fibra penetra nei rivestimenti del muscolo e nel perimisio dà luogo ad un certo numero di ramificazioni Queste originano a livello di un nodo di Ranvier e ciascuna ramificazione mantiene entrambe le guaine Ciascuna di queste ramificazioni stabilisce un contatto sinaptico con una fibra muscolare diversa il contatto sinaptico si chiama placca motrice o sinapsi neuromuscolare Ciascun ramo della fibra nervosa originaria raggiunge una fibra muscolare dà luogo ad una serie di ramificazioni sottili che costituiscono nel complesso quella che viene definita arborizzazione terminale A questo livello il ramo della fibra nervosa perde la guaina mielinica ma mantiene un rivestimento analogo alla guaina di Schwann formata dalle cellule di teloglia o cellule di Schwann terminali che formano una specie di involucro intorno all’arborizzazione. Questo involucro somiglia ad una campana la cui base prende contatto con la fibra muscolare Si possono osservare i nuclei delle cellule di teloglia che appaiono di forma allungata e vengono chiamati nuclei della arborizzazione terminale Con il cloruro d’oro si mette in evidenza tutto il complesso di rami dell’arborizzazione terminale, rami che prendono contatto con il sarcolemma della fibra muscolare e questo rapporto di contiguità prende il nome di placca motrice o giunzione o sinapsi neuromuscolare UNITÀ MOTORIA Per unità motoria si intende l’insieme delle fibre muscolari che vengono controllate da un singolo motoneurone. Il numero delle fibre di una unità motoria può variare fino a raggiungere qualche centinaia esiste un rapporto inverso di questo numero rispetto al grado di specializzazione muscolare nei muscoli estrinseci dell’occhio controllare una sola fibra muscolare un motoneurone può nei muscoli soggetti a movimenti meno delicati (es. gluteo) un motoneurone controlla alcune centinaia di fibre ELEMENTI DELLA SINAPSI NEUROMUSCOLARE STRUTTURA PRESINAPTICA I rami della arborizzazione terminale svolgono un ruolo di struttura presinaptica mentre il sarcolemma e le strutture che si trovano all’interno di esso diventano la struttura postsinaptica A livello della arborizzazione terminale si trovano: – mitocondri – alcuni costituenti del citoscheletro (es. microtubuli) – vescicole sinaptiche (del diametro di 50-60nm) che contengono il mediatore chimico tipico di questa sinapsi, rappresentato dall’acetilcolina I rami della arborizzazione terminale della placca motrice, a differenza delle sinapsi neuronali o centrali, non terminano con bottoni terminali in tutti i casi, ma spesso tendono ad assottigliarsi alle loro estremità l’assolemma che riveste questi rami svolge quindi il ruolo di membrana presinaptica 7 TESSUTO MUSCOLARE SCHELETRICO STRUTTURA POSTSINAPTICA la fibra muscolare, nella zona del contatto sinaptico, presenta un lieve sollevamento del sarcolemma (SUOLA SUOLA DELLA PLACCA MOTRICE) MOTRICE dovuto a: •Maggiore presenza di sarcoplasma dove abbondano ribosomi e mitocondri •Numero più elevato di nuclei appartenenti al sincizio della fibra muscolare. Questi nuclei vengono detti nuclei della suola della placca motrice Il sarcolemma svolge il ruolo di membrana postsinaptica, si adatta ai rami terminali dando luogo a invaginazioni che rappresentano le impronte dei rami stessi Queste invaginazioni si chiamano docce o fessure sinaptiche primarie Sul fondo di ciascuna doccia si trovano ulteriori invaginazioni, dette docce o fessure sinaptiche secondarie che nel complesso costituiscono l’apparato subneurale L’assolemma si adatta alla doccia primaria mentre non penetra all’interno delle docce secondarie VALLO SINAPTICO Tra la membrana presinaptica e la membrana postsinaptica si trova uno spazio di circa 40nm che viene denominato spazio o vallo sinaptico al suo interno si trova del materiale glicoproteico che al microscopio elettronico ha un aspetto elettrondenso fioccoso contenente un enzima capace di degradare l’ acetilcolina, (acetilcolinaesterasi) il materiale proteico, al contrario dell’assolemma, si insinua anche nelle docce secondarie a livello del sarcolemma si trovano i recettori per l’acetilcolina si tratta di complessi proteici disposti a costituire un canale ionico essi contengono delle unità monomeriche di cui una ha la capacità di legare il mediatore chimico per quanto riguarda il meccanismo di trasmissione sinaptico, esso non differisce da quello della sinapsi centrale IL MECCANISMO DELLA CONTRAZIONE lo stimolo per la contrazione raggiunge il sarcolemma tramite la placca motrice lungo il sarcolemma viene propagato ai centrotubuli. a loro volta i centrotubuli possono trasmettere il segnale nervoso alle cisterne terminali. queste rispondono con la liberazione di ioni calcio che vengono a trovarsi liberi nel sarcoplasma. Gli ioni Ca++ interagisono con la subunità C della troponina del filamento sottile, provocando un cambiamento dei rapporti tra le varie subunità della troponina (le subunità si avvicinano tra di loro rinsaldando i legami reciproci), a seguito di ciò cambiano anche i rapporti tra la tropomiosina e l’actina globulare la troponina I non maschera più i siti di legame dell’actina globulare per la miosina i ponti trasversi della miosina possono legarsi all’actina globulare l’ATPasi della subunità S1 della miosina scinde l’ATP presente nella stessa subunità e da luogo ad ADP + Pi + E l’energia fornita dalla scissione dell’ATP consente una oscillazione del ponte trasverso (zone di flessione tra S1 e S2 e tra S2 e MML) l’oscillazione della miosina (subunità S1 o ponte trasverso) trascina il filamento di actina verso il centro del sarcomero in misura di circa 100A° l’arrivo di una nuova molecola di ATP provoca il distacco della subunità S1dall’actina globulare (in assenza di ATP il distacco non avviene: complessi rigor) se permane lo stimolo ed è presente ATP, si ripetono cicli di attacco, oscillazione, distacco ecc. con accorciamento di tutti i sarcomeri, miofibrille, fibre di un determinato muscolo 8 TESSUTO MUSCOLARE SCHELETRICO la miosina, con le oscillazioni dei ponti trasversi, trascina i filamenti sottili verso il centro del sarcomero. in definitiva si ha l’accorciamento del sarcomero dovuto a scorrimento dei filamenti sottili verso il centro del sarcomero trascinati dal movimento dei ponti trasversi. nel sarcomero contratto scompaiono gli emidischi I e la banda H Tessuto Muscolare Striato Scheletrico Contrazione RILASCIAMENTO DEL MUSCOLO Quando termina lo stimolo per la contrazione e la sinapsi neuromuscolare cessa la trasmissione dell’impulso al sarcolemma: il calcio viene riportato all’interno del reticolo sarcoplasmatico perché entrano in azione le pompe del Ca++ ATP dipendenti presenti nelle cisterne terminali; le subunità di troponina tornano all’assetto delle condizioni di riposo (legami più lassi); i siti dell’actina globulare vengono di nuovo mascherati; i ponti trasversi, dopo l’ultimo ciclo di oscillazione, a seguito del distacco dall’actina globulare provocato dall’ATP, non possono più agganciarsi all’actina globulare. A questo punto prevalgono le forze del rilasciamento muscolare: azione dei muscoli antagonisti, trazione esercitata dalle inserzioni tendinee, i sarcomeri tornano nello stato di riposo, compaiono di nuovo gli emidischi I e banda H. 9 TESSUTO MUSCOLARE SCHELETRICO IL TESSUTO MUSCOLARE DEL MIOCARDIO FORMA DEI CARDIOMIOCITI E COMPORTAMENTO NELLA •costituisce la componente contrattile del cuore •le cellule che lo costituiscono sono dette cardiomiociti o miociti cardiaci •si tratta di cellule dotate ciascuna della propria autonomia •a differenza del muscolo scheletrico i miociti non danno luogo alla formazione di sincizi •sono strettamente correlate tra di loro sia mediante i nexus che da altre specializzazioni giunzionali •i nexus permettono alle cellule di interagire funzionalmente in maniera molto coordinata •per questo motivo si dice che le cellule cardiache costituiscono un sincizio funzionale COSTITUZIONE DEL MIOCARDIO MORFOLOGIA AL MICROSCOPIO OTTICO IN SEZIONE TRASVERSALE • • • in una sezione trasversale si possono osservare dei contorni piuttosto irregolari delle cellule si trovano sezioni di calibro ristretto accanto a sezioni di calibro maggiore senza che vi sia una gradualità nelle dimensioni le sezioni più piccole corrispondono ai prolungamenti mentre quelle più grandi alle parti centrali delle cellule: infatti nella maggior parte dei casi le sezioni più grandi contengono il nucleo (grande e in posizione centrale) • i cardiomiociti presentano una forma grossolanamente cilindrica nella loro porzione centrale • alle estremità ogni cellula presenta dei prolungamenti piuttosto tozzi che si connettono con prolungamenti analoghi di altre cellule • in altre parole i cardiomiociti si anastomizzano tra di loro in posizione termino-terminale • da ciò deriva che le superfici laterali difficilmente stabiliscono contatti con le cellule adiacenti ma in genere delimitano degli spazi che vengono occupati da connettivo piuttosto lasso ricco di vasi MORFOLOGIA AL MICROSCOPIO OTTICO IN SEZIONE LONGITUDINALE • in una sezione longitudinale si nota che le cellule del miocardio non presentano una striatura trasversale così evidente come quella del muscolo scheletrico –ciò dipende da diversi fattori: •i miofilamenti pur essendo organizzati a livello molecolare come nel muscolo scheletrico costituiscono fasci ma non danno mai luogo a grosse miofibrille •nelle cellule cardiache sono più abbondanti il sarcoplasma e i mitocondri. In conseguenza di ciò i fasci di filamenti contrattili sono distanziati tra di loro e non risultano perfettamente in fase come avviene per le miofibrille del muscolo scheletrico • un’ altro elemento caratteristico nella sezione longitudinale del muscolo cardiaco è rappresentato dalla struttura che si presenta in prossimità dei confini tra cellula e cellula, questa struttura viene detta stria scalariforme o disco intercalare 10 TESSUTO MUSCOLARE SCHELETRICO DISCHI INTERCALARI O STRIE SCALARIFORMI •l’aspetto dei dischi intercalari deriva dal fatto che quando due cellule cardiache si mettono in rapporto tra di loro in posizione termino-terminale formano dei confini a livello dei quali le membrane possono decorrere in due maniere: –in senso trasversale, trasversale obliquo o perpendicolare rispetto all’ asse maggiore della cellula –in senso longitudinale, longitudinale cioè parallelo all’asse maggiore della cellula TUBULI T E RETICOLO SARCOPLASMATICO I centrotubuli sono analoghi a quelli del muscolo scheletrico; sono costituiti da invaginazioni del sarcolemma e non si trovano al confine tra emidisco chiaro e disco scuro ma in prossimità della linea Z i centrotubuli del miocardio hanno un calibro maggiore rispetto a quelli del muscolo scheletrico la superficie interna dei tubuli T è ricoperta da materiale glicoproteico che corrisponde al materiale che si trova sul sarcolemma stesso il reticolo sarcoplasmatico è diverso da quello del muscolo scheletrico: non sono presenti cisterne terminali e fenestrate i sarcotubuli, giunti in prossimità dei centrotubuli si slargano leggermente formando dei pedicelli che stabiliscono contatti con i centrotubuli analoghi rispetto a quelli delle triadi del muscolo scheletrico non si può parlare di triadi e invece questi rapporti sarcotubuli-centrotubuli sono detti diadi. tratti trasversali –assumono una maggiore densità anche al microscopio ottico e per questo motivo i confini tra cellula e cellula appaiono come degli scalini e da qui il loro nome (strie scalariformi) –al microscopio elettronico si è visto che in questi tratti le membrane adiacenti danno luogo a numerose escrescenze che si interdigitano tra di loro e ciò è motivo di maggiore densità –questi tratti contengono numerose specializzazioni come desmosomi, nexus e zonulae adherentes (non essendo perimetrali invece di zonulae si chiamano più correttamente fasciae adherens) •tratti longitudinali –mancano le interdigitazioni delle membrane che così decorrono linearmente parallele tra di loro –contengono specializzazioni del tipo nexus mentre sono assenti altre specializzazioni IL TESSUTO MUSCOLARE LISCIO Localizzazione: • si trova largamente rappresentato nell’organismo infatti costituisce: – parete muscolare del tubo digerente, dall’esofago (esclusa la prima porzione) fino all’intestino crasso – pareti delle arterie – muscoli intrinseci dell’occhio – pareti degli ureteri e della vescica – vie respiratorie (si trova nella trachea, nei bronchi fino nelle loro ramificazioni di calibro più piccolo) – utero e tube uterine – un particolare tipo di cellule muscolari lisce si trova intorno agli adenomeri di alcune ghiandole, queste cellule capaci di attività contrattili si chiamano mioepiteliali Origine: • le cellule muscolari derivano da cellule mesenchimatiche indifferenziate che seguono un destino diverso da quello connettivale • fanno eccezione: • le cellule della muscolatura intrinseca dell’occhio nella quale le cellule hanno origine ectodermica • le cellule mioepiteliali che possono originare dai foglietti ectodermico ed entodermico 11 TESSUTO MUSCOLARE SCHELETRICO DIMENSIONI E FORMA • • • • • le cellule muscolari lisce hanno aspetto allungato e per questo vengono dette fibrocellule ciascuna cellula ha una forma affusata con una parte centrale più larga contenente il nucleo le fibrocellule possono avere una lunghezza variabile – minimo 20 - 40 micron, nei muscoli intrinseci dell’occhio e nella parete delle piccole arterie – massimo 500 micron nell'utero gravido si possono trovare in numero esiguo ma spesso formano dei veri e propri strati o tonache muscolari all’interno delle tonache muscolari le cellule si dispongono parallelamente tra di loro ma sfasate in maniera che l’estremità di una cellula viene compresa tra l’estremità di altre due cellule MORFOLOGIA AL MICROSCOPIO OTTICO sezione longitudinale • le fibrocellule appaiono distanziate di circa 60-90 nm (a volte anche 100 nm) • nei punti dove le cellule stabiliscono nexus la distanza è ridotta a 2 nm • gli spazi intercellulari sono occupati da tessuto connettivo reticolare; può essere evidenziato con il metodo del PAS o con impregnazione argentica; forma una rete tridimensionale che impedisce alle cellule di allontanarsi durante la contrazione. • ciascuna fibrocellula, mostra il nucleo nella zona centrale • in genere il nucleo copre una lunghezza pari al 10% dell’intera lunghezza della fibrocellula • il nucleo ha un aspetto piuttosto allungato che ripete la forma della fibrocellula MORFOLOGIA AL MICROSCOPIO ELETTRONICO A TRASMISSIONE sezione trasversale • il tessuto muscolare liscio appare costituito da una serie di poligoni abbastanza regolari che corrispondono alle fibrocellule interessate dal piano della sezione • le sezioni cellulari di forma poligonale hanno dimensioni variabili • è presente una certa gradualità, sezioni più piccole corrispondono all’estremità della cellula, sezioni più grandi in genere contengono il nucleo • il nucleo, quando è presente nella sezione, è localizzato in posizione eccentrica a differenza di quello che accade nel muscolo scheletrico dove i nuclei sono più di uno e periferici Lungo il contorno del sarcolemma si notano piccole insenature dette caveole • Esiste una relazione tra la presenza di queste caveole e la trasmissione dell’impulso nervoso e l’avvio del meccanismo contrattile (similitudine funzionale con i centrotubuli) • all’interno delle fibrocellule sono stati riscontrati tre tipi di filamenti: – microfilamenti di actina (5-6 nm Ø) – filamenti intermedi (10nm Ø) di desmina, vimentina, filamina e sinemina – filamenti contrattili (15 nm Ø) di miosina; non sono facilmente dimostrabili come nel muscolo scheletrico (probabilmente si assemblano e si disassemblano) 12 TESSUTO MUSCOLARE SCHELETRICO MECCANISMO DI CONTRAZIONE DELLE FIBROCELLULE • • • 1. 2. 3. 4. 5. 6. all’interno del citoplasma della fibrocellula si trovano dei corpi elettrondensi costituiti da alfa actinina, sono detti corpi densi citoplasmatici sul versante interno del sarcolemma si trovano formazioni simili che vengono dette placche dense subsarcolemmali (contengono proteine di ancoraggio come talina e vinculina) il sarcoplasma contiene inoltre: – mitocondri, reticolo endoplasmatico liscio, (disseminati ovunque) – ribosomi, apparato del Golgi, 2 centrioli, (a ciascun polo del nucleo in una regione di forma conica) – gocce lipidiche e di glicogeno, microtubuli quando si verificano le condizioni per la contrazione, l’impulso nervoso raggiunge il sarcolemma, lo percorre e, per mezzo delle caveole, si porterebbe in profondità nella cellula dove sono presenti strutture del reticolo endoplasmatico liscio ciò determinerebbe il rilascio di ioni Ca++ dal reticolo liscio gli ioni Ca++ si legano prontamente alla calmodulina, una proteina che innesca il meccanismo contrattile. (nel muscolo liscio non si trova la troponina C) il complesso Ca-Calmodulina si lega a una proteina chinasi (chinasi della catena leggera della miosina) che determina interazione actina miosina vengono attivati i sistemi ATPasici della miosina cosicchè i filamenti di miosina interagiscono con quelli di actina probabilmente in maniera simile a quella che avviene nel muscolo scheletrico (scorrimento reciproco) in conseguenza di ciò le placche subsarcolemmali vengono avvicinate ai corpi densi con il risultato di determinare un accorciamento della cellula • • • sembra probabile che i filamenti di tipo intermedio colleghino i corpi densi tra di loro e i corpi densi alle placche subsarcolemmali formerebbero così un reticolo che fa da sostegno ai filamenti di actina e di miosina i microfilamenti di actina e i filamenti di miosina formerebbero dei fascetti analoghi alle miofiofibrille (senza striatura) tesi tra i corpi densi e le placche subsarcolemmali CLASSIFICAZIONE FUNZIONALE DEL TESSUTO MUSCOLARE LISCIO nell’ambito del nostro organismo si distinguono due tipi di tessuto muscolare liscio: • multiunitario, muscoli intrinseci dell’occhio e nei piccoli vasi • viscerale: si trova nel tubo digerente, nella parete dei grossi vasi, nell’ utero • nel tipo multiunitario ciascuna cellula è dotata di una propria terminazione nervosa (proveniente dal sistema nervoso autonomo) • nel tipo viscerale solo alcune cellule hanno una terminazione nervosa – si ritiene che l’ impulso possa passare dalle cellule dotate di terminazione a quelle che ne sono prive per mezzo di nexus che infatti sono molto abbondanti in questo tipo 5 4 2 3 13 TESSUTO MUSCOLARE SCHELETRICO ULTRASTRUTTURA DEL NEXUS NEXUS O GAP JUNCTION • • • • • • denominato anche gap junction, giunzione comunicante, facilitante o sinapsi elettrica si riteneva che fosse presente solo in alcuni tessuti, invece è diffuso a tutti i tessuti dell'organismo compresi i tessuti embrionali la microscopia elettronica a trasmissione mise in evidenza che in alcuni tratti dei confini tra cellule adiacenti le membrane cellulari potevano avvicinarsi ad una distanza di 2 nm l'impiego di traccianti per la microscopia elettronica come il lantanio o il rosso rutenio dimostrò che in queste zone di avvicinamento delle due membrane erano comunque presenti degli spazi intervallati da punti di connessione fra cellula e cellula questi punti di connessione erano costituiti da protuberanze poligonali a forma esagonale denominati connessoni il nexus quindi è costituito da: – un’area della membrana citoplasmatica di forma rotondeggiante avente un diametro che può anche raggiungere circa 5µ – quest’area è punteggiata da connessoni disposti regolarmente – sulla membrana della cellula adiacente si trova un area corrispondente • • • • • • • • mediante la tecnica del criodecappaggio si e visto che ciascun connessone ha il diametro di circa 7 nm e la distanza dei connessoni vicini, da centro a centro, è di circa 10 nm ciascun connessone é a sua volta costituito da 6 subunità proteiche formate da una proteina detta connessina (isolate varie isoforme con26, con 31, ecc) queste sei subunità si dispongono attorno ad un canalicolo centrale avente diametro medio di 1,5 – 2 nm le singole unità di connessina (quindi il connessone nel suo complesso) sono proteine transmembranali che si portano dall'interno del citoplasma all'estemo del doppio strato lipidico sul versante esterno il connessone protrude di circa 0,9-1 nm rispetto al piano del doppio strato lipidico nel costituire il nexus i connessoni che sporgono su una membrana si appoggiano ai connessoni che sporgono dalla membrana adiacente il canalicolo di un connessone si continua con quello dell'altro e alla fine mette in comunicazione l'interno di una cellula con l'interno dell'altra la sporgenza di ciascun connessone di 0,9-1 nm sui due doppi strati delle cellule adiacenti fa si che le due cellule risultino distanziate di 1,8-2 nm FUNZIONI DEL NEXUS • Permette il passaggio di sostanze dal citoplasma di una cellula a quello della cellula immediatamente vicina • infatti la disposizione delle subunità di connessina è leggermente obliqua e tale da poter determinare apertura o chiusura del canalicolo del connessone come il diaframma della macchina fotografica • l’apertura avviene in varie condizioni: – abbassamento del pH – aumento di Ca++ libero a seconda di quali sostanze attraversano il nexus si può avere: avere: • accoppiamento elettrico fra due cellule – sono zone a bassa resistenza elettrica – permettono il passaggio di ioni Na+ il chè consente la trasmissione del potenziale d'azione da una cellula all'altra • accoppiamento funzionale o metabolico tra due cellule – possono attraversare il nexus molecole a basso peso molecolare (<1000 Daltons) come ad es. i secondi messaggeri • AMP ciclico • GMP ciclico – determinano accoppiamento funzionale anche: • ioni Ca++ • ioni Mg++ • altro… 14
Scaricare