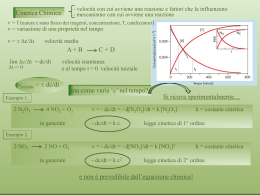
Cinetica Chimica ¾ La Cinetica Chimica descrive l’andamento nel tempo della reazione che partendo dai reagenti forma i prodotti Reagenti → Prodotti ¾ Il rateo di reazione (r) è definito come la velocità di trasformazione di ciascun componente per unità di volume d [x ] r= ⇒ dt E.Giusti: Cinetica Batterica R =V ×r con [x] = concentrazione pag. 1 Ordine della cinetica ¾ E’ data dalla somma degli esponenti di reazione (determinati sperimentalmente) ¾ Ordine zero: velocità di reazione costante, indipendente dalla concentrazione dA del reagente [A] → Prodotti = − ko dt ¾ Ordine uno: velocità di reazione proporzionale alla concentrazione [A] → Prodotti dA = − ko A dt ¾ Ordine due: velocità proporzionale al quadrato della concentrazione o al prodotto di due concentrazioni dA 2 2 [A] → Prodotti [A] + [B] → Prodotti E.Giusti: Cinetica Batterica dt = − ko A dA dB = = − k o A⋅ B dt dt pag. 2 Cinetica del primo ordine 1 0.9 C( t ) = e 0.8 − k ⋅t La tangente alla curva per t=0 intercetta l’asse dei tempi per un valore pari a 1/k 0.7 0.6 C(t) 0.3679 −k 0.5 Al quell’istante, la concentrazione si è ridotta a 1/e della concentrazione iniziale 0.4 0.3 0.2 1 k 0.1 0 0 1 E.Giusti: Cinetica Batterica 2 3 4 5 6 7 8 9 10 pag. 3 Cinetica Enzimatica La trasformazione da reagenti a prodotti è mediata da un enzima che ha la funzione di combinarsi temporaneamente con il reagente (substrato) formando un complesso attivato instabile (ES*), che poi forma il prodotto finale (P). Il meccanismo di attivazione dell'enzima (E) consiste spesso nel collocarsi in una particolare posizione del substrato (in genere costituito da complesse catene proteiche) in modo da catalizzare la trasformazione del substrato (S) in Prodotto (P). SUBSTRATO COMPLESSO ATTIVATO E + S → ES* → E + P ENZIMA LIBERO Riutilizzo dell’enzima E.Giusti: Cinetica Batterica pag. 4 Cinetica di Michaelis-Menten μ (S ) = μ max Ks + S S La cinetica dipende in modo nonlineare dalla concentrazione del substrato S Inoltre essa dipende da due parametri: • µmax rappresenta la massima velocità di reazione • Ks esprime la dipendenza dall’enzima E.Giusti: Cinetica Batterica per alti valori del substrato la cinetica approssima l’ordine zero μ( S ) ( 1 / h ) μ max 0.5 K s = 5 mg/l 0.45 0.4 μ max 0.35 K s = 25 mg/l K s = 50 mg/l Ks K s = 100 mg/l 0.3 1 μ max 2 0.25 0.2 0.15 0.1 0.05 0 05 per bassi valori del substrato la cinetica tende al primo ordine con velocità 25 50 S (mg/l) 100 150 Ks è detta costante di mezza velocità perché quando S = Ks la velocità di reazione è la metà del massimo pag. 5 Cinetica a più substrati limitanti ¾ La cinetica può dipendere dalla disponibilità contemporanea di più substrati ¾ Ciascuno dei substrati è detto limitante, perché la sua concentrazione modula la cinetica LEGGE DI LIEBIG: Il substrato più scarso determina la velocità della cinetica S1 S2 Sn μ (S1 ,S2 ,...,Sn ) = μ max × × ×...× K1 + S1 K2 + S2 Kn + Sn E.Giusti: Cinetica Batterica pag. 6 Legge di Liebig . μ(S1 ,S2 ) = μmax Limitazione del primo substrato S1 S2 ⋅ Ks1 +S1 Ks2 +S2 Limitazione del secondo substrato 0.35 0.3 Cinetica (1/h) 0.25 0.2 0.15 0.1 0.05 0 50 40 50 30 Substrato 2 (mg/l) Ks2 = 25 mg/ l E.Giusti: Cinetica Batterica 40 30 20 20 10 10 0 0 Substrato 1 (mg/l) Ks1 = 5 mg/ l pag. 7 Effetto di un limitatore esterno μ ( S1 , S 2 ) = μ max In altri casi, si può avere un elemento, diverso dal substrato, che limita la cinetica inversamente alla sua concentrazione. K s2 S1 ⋅ K s1 + S1 K s2 + S 2 Rateo cinetico (1/h) 0.5 Non è propriamente un inibitore, perché non blocca la complessazione dell’enzima, ma rallenta la cinetica con legge iperbolica. 0.4 0.3 0.2 0.1 0 60 S1 40 20 Substrato limitante (mg/l) Ks1 = 1 5 mg/ l E.Giusti: Cinetica Batterica 0 10 8 6 4 2 S2 0 La cinetica rimane monotona, con µmax che varia con legge iperbolica rispetto a questo elemento Limitatore esterno Ks2 = 1 mg/l pag. 8 Cinetica di Monod ¾ Negli anni ‘30 - ‘40 Monod iniziò a studiare la cinetica dei batteri ¾ Cercò di determinare un legame fra concentrazione di substrato limitante e velocità di crescita ¾ Per i suoi esperimenti usò un batterio estremamente diffuso e conosciuto: l’ Escherichia coli ¾ Come substrato usò Acido lattico ¾ Osservò sperimentalmente che la velocità di accrescimento seguiva un andamento analogo a quello che Michaelis-Menten avevano trovato per le reazioni enzimatiche ¾ Perciò concluse che l’accrescimento batterico avveniva attraverso reazioni enzimatiche E.Giusti: Cinetica Batterica pag. 9 Evoluzione di una popolazione batterica in un batch Si insemina il batch con una quantità iniziale ¾ Si osservano quattro fasi: di biomassa (inoculo) e si fornisce una ¾ Incubazione: la biomassa si deve quantità iniziale di substrato. acclimatare al nutriente presente e la sua crescita è quasi nulla Non si aggiunge altro durante l’esperimento ¾ Crescita esponenziale: una volta Si osserva come variano le concentrazioni di adattata al substrato, la biomassa substrato e di biomassa cresce con ritmo proporzionale alla sua concentrazione Substrato ¾ Stasi: la crescita raggiunge un livello per il quale il substrato rimasto non è sufficiente a sostenere la crescita Biomassa ¾ Decadimento: mancando il substrato, nella biomassa prevale il metabolismo endogeno, con tempo consumo di sostanza ossidabile crescita decadimento incubazione esponenziale stasi all’interno della biomassa, fino alla sua estinzione E.Giusti: Cinetica Batterica pag. 10 Cinetica di Monod ¾ Crescita batterica basata sulle seguenti ipotesi ¾ ¾ ¾ ¾ L’enzima è proporzionale alla biomassa La sintesi batterica è proporzionale al consumo di substrato Si modella anche il metabolismo endogeno, proporzionale alla biomassa Si introduce il fattore di resa Y = biomassa prodotta/substrato consumato Dinamica del substrato Dinamica della biomassa dS 1 μ max S =− X dt Y Ks + S μ max S dX = X − bH X dt Ks + S Enzima proporzionale alla biomassa μ max X Decadimento proporzionale alla biomassa −bH X Fattore di resa (Yield factor) E.Giusti: Cinetica Batterica Y= ΔX sintetizzata ΔS utilizzato pag. 11 Varianti metaboliche Growth-decay: Il metabolismo endogeno produce solamente massa inerte Xi, che però il modello non descrive (approccio classico e ASM1) Death-regeneration: Il metabolismo endogeno produce in parte (fP) massa inerte Xi ed in parte (1-fP) nuovo substrato in conseguenza della lisi cellulare. Il modello genera la parte nonbiodegradabile del substrato SI (approccio ASM3) E.Giusti: Cinetica Batterica XH growth bH X H XI decay SS Substrato esterno f P bH X H Frazione mineralizzata della biomassa decaduta XH XI death SS regeneration hydrolysis XS (1 − f P )bH X H Frazione biodegradabile della biomassa decaduta Substrato esterno pag. 12 Modelli di metabolismo endogeno ¾ Modello Growth-decay (classico): La biomassa decade producendo solamente sostanze inerti (mineralizzazione) Ss dS s 1 XH = − μ max dt Y K s + Ss Ss dX H X H − bH X H = μ max dt K s + Ss ¾ Modello Death- regeneration (ASM3): Il decadimento della biomassa produce nuovo substrato per la parte non mineralizzata (1 − f P ) Ss dS s 1 X H + (1 − f P )bH X H = − μ max dt Y K s + Ss Ss dX H X H − bH X H = μ max dt K s + Ss E.Giusti: Cinetica Batterica pag. 13 L’approccio ASM3 tiene conto della mineralizzazione 20 Substrato 18 14 mgCOD L-1 Si ha la scomparsa totale del substrato. Ciò significa che esso è supposto completamente biodegradabile e non viene prodotto alcun residuo mineralizzato (non biodegradabile) Modello classico Growth - decay 16 12 10 Biomassa 8 6 4 2 0 0 5 10 15 20 20 30 35 40 45 50 Substrato 18 Modello ASM3 Death - regeneration 16 14 mgCOD L-1 25 tempo 12 10 dS s Ss 1 X H + (1 − f P )bH X H → 0 = − μ max dt Y K s + Ss Biomassa 8 6 4 S I = 0.4616 2 0 0 5 10 15 E.Giusti: Cinetica Batterica 20 25 tempo 30 Solamente la frazione (1 - fP) di substrato viene metabolizzata, mentre la rimanente parte si converte in substrato NON biodegradabile (mineralizzazione). Il suo valore (SI) si ricava azzerando la derivata del substrato. 35 40 45 50 SI = K s (1 − f P )bH μ max Y − (1 − f P )bH pag. 14 Passaggio da sintesi a endogeno S* = SI K s bH μ max − bH S Evoluzione nel tempo 8 X = X max Evoluzione nel piano di fase S, X 7 X = X max X -1 Biomassa (mg COD L) 6 5 S = S* S = SI 4 3 2 S o( 1 ) = 5 1 Biomassa iniziale Xo 0 0 2 E.Giusti: Cinetica Batterica 4 S o( 2 ) = 10 6 8 10 12 -1 Substrato (mg COD L) S o( 3 ) = 15 14 16 S o( 4 ) = 20 18 20 Il massimo valore raggiunto dalla biomassa Xmax dipende dalle condizioni iniziali. In ogni caso esso corrisponde al valore di substrato S*. Tutte le traiettorie hanno SI per valore asintitotico pag. 15 Cinetiche più utilizzate nei modelli Cinetica di Monod rM = μmax Assimilazione diretta di substrato solubile e trasformazione in biomassa Ss rM XH Cinetica endogena Perdita di biomassa e trasformazione parziale (1-fp) in nuovo substrato per lisi cellulare. La rimanente quota fp viene mineralizzata (1 − f p ) XH fp X s ,Ss Trasformazione di materiale organico particolato in solubile, ad opera di batteri eterotrofi Xs E.Giusti: Cinetica Batterica Ss re = −bH X H death re = 1 − f p bH X H regeneration ( ) XI Cinetica di idrolisi rh Ss XH K s + Ss X s /X H rh = k h XH K X + X s /X H Il rateo dipende dal rapporto fra particolato (Xs) e biomassa (XH) pag. 16 Modelli cinetici complessi ¾ La biomassa eterotrofa può svolgere molti processi contemporaneamente ed operare su substrati con diverse caratteristiche di degradabilità. ¾ La catena di processi più completa è contenuta nei modelli ASM ¾ Idrolisi del substrato particolato ¾ Fermentazione con produzione di acetato ¾ Crescita aerobica su substrato fermentabile ¾ Crescita aerobica su substrato fermentato (acetato) ¾ Denitrificazione su substrato fermentabile ¾ Denitrificazione su substrato fermentato (acetato) ¾ Lisi cellulare (decadimento) ¾ Ogni rateo cinetico è del tipo Monod a substrato multiplo, eventualmente con termini modulanti (limitatore esterno) r = μ max S Ko × × ....... × X biomassa K s + S K o + So substrato lim i tan te E.Giusti: Cinetica Batterica lim itatore esterno pag. 17 Rappresentazione matriciale: eq. di rateo ¾ I fattori che compongono il modello sono: ¾ ¾ ¾ ¾ Componenti del processo i Processi di trasformazione j Coefficienti stechiometrici (equivalenze in peso) νji Cinetiche (strutturazione delle reazioni chimiche) ρj ¾ Il rateo di produzione della componente i, ri, si ottiene come somma di tutti i processi paralleli ρj che comprendono la variabile i, attraverso i coefficienti stechiometrici νji ri = ∑ v ji ρ j j ¾ Dei vari coefficienti stechiometrici νji uno può essere posto adimensionalmente pari a ± 1, mentre per tutti gli altri devono essere definite delle equazioni algebriche che tengono conto della conservazione della massa E.Giusti: Cinetica Batterica pag. 18 Rappresentazione matriciale: eq. di continuità ¾ Rendono conto del principio che nelle reazioni chimiche gli elementi, elettroni (o COD) e cariche elettriche non vengono né formate né distrutte, ma solo trasformate ¾ La stechiometria del modello è basata su tre principi di continuità per ¾ ¾ ¾ ¾ COD cariche elettriche Azoto Fosforo ¾ L’equazione generica di continuità, valida per tutti i processi j e per tutte i materiali c soggetti a continuità è 0 = ∑ vij ici ∀i j ¾ I fattori ici convertono le unità del componente i nelle unità del materiale c, al quale deve essere applicata l’eq. di continuità. E.Giusti: Cinetica Batterica pag. 19 Rappresentazione matriciale Componente Processo Cinetica S1 S2 r1 v11 v12 ρ1 r2 v21 v22 ρ2 ri = ∑ vij ρ j j E.Giusti: Cinetica Batterica dS1 = r1 = v11 ρ 1 + v21 ρ 2 dt dS 2 = r2 = v12 ρ 1 + v22 ρ 2 dt pag. 20 Rappresentazione matriciale Assimilazione di un substrato biodegradabile da parte di biomassa eterotrofa (senza metabolismo endogeno) Ss substrato XH Processo Crescita eterotrofa Substrato (SS) − Biomassa (XH) 1 YH dS S SS 1 =− μ max XH dt YH K s + SS SS dX H = μ max XH dt K s + SS E.Giusti: Cinetica Batterica biomassa 1 Cinetica μ max SS XH K s + SS La dinamica di ciascun componente si ottiene moltiplicando la cinetica per il corrispondente fattore 1 di resa o 1 YH pag. 21 Rappresentazione matriciale (2) Assimilazione di un substrato biodegradabile da parte di biomassa eterotrofa (con metabolismo endogeno) Ss substrato XH Processo Crescita eterotrofa Metabolismo endogeno Substrato (SS) − 1 YH 0 (1 − f P ) biomassa Biomassa (XH) 1 −1 Cinetica μ max SS XH K s + SS bH X H dS S SS 1 =− X H + (1 − f P )bH X H μ max dt YH K s + SS SS dX H = μ max X H − bH X H dt K s + SS E.Giusti: Cinetica Batterica pag. 22 Rappresentazione matriciale del modello ASM1 j↓ i→ Biomassa XB Growth 1 Decay −1 ri = Ratei Unità Substrato SS 1 − Y mgCOD/L Ossigeno So 1−Y − Y Processi ρ Ss ˆμ XB K s + Ss −1 bH X B ∑j vij ⋅ ρ j mgCOD/L mgCOD/L Ss dS s 1 = − μˆ XB dt Y K s + Ss Ss dX B X B − bH X B = μˆ dt K s + Ss dS o Ss 1−Y μˆ X B − bH X B =− dt Y K s + Ss E.Giusti: Cinetica Batterica pag. 23 Bibliografia Franks R.G.E., Modelling and Simulation in Chemical Engineering, Wiley, 1972. Carrà S. e Forni L. , Aspetti Cinetici della Teoria del Reattore Chimico, Tamburini editore, Bologna, 1974. Gavezzotti A., Cinetica Chimica, Guadagni editore, Milano, 1982. Bungay, H.R., Computer Games and Simulation for Biochemical Engineering, 1985. Bailey J.E. e Ollis D.F. Biochemical Engineering Fundamentals, McGraw-Hill, 1986. Marsili-Libelli, S. “Modelling, Identification and Control of the Activated Sludge Process”, Advances in Biochemical Engineering/Biotechnology (A. Fiecther ed.), vol. 38: 90 - 148, 1989. Walas, S.M. Modelling with Differential Equations in Chemical Engineering, Butterworth, 1991. Orhon D., Artan N., Modelling of Activated Sludge Systems, Technomic Publ. Co., 1994. E.Giusti: Cinetica Batterica pag. 24
Scaricare