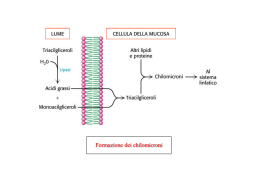
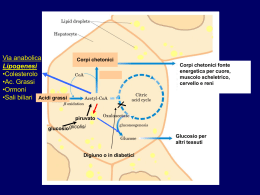
Energia di ossidazione Kcal/g Kj/g Grassi 9 38 Carboidrati 4 17 Proteine 4 17 Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine I grassi della dieta vengono assorbiti nell’intestino tenue Struttura molecolare di un chilomicrone Formazione dei chilomicroni L’utilizzo degli acidi grassi come combustibili richiede tre stadi di trasformazione 1) I lipidi devono essere mobilizzati. In questo processo i triacilgliceroli vengono degradati ad acidi grassi e glicerolo, che vengono rilasciati dal tessuto adiposo e trasportati ai tessuti che richiedono energia. 2) Gli acidi grassi devono quindi essere attivati e trasportati nei mitocondri per la degradazione. 3) Nei mitocondri gli acidi grassi vengono degradati in modo graduale in acetil CoA, che poi viene ossidato nel ciclo dell’acido citrico. Il primo passaggio nell’utilizzazione dei grassi come fonte di energia è l’idrolisi dei trigliceridi da parte di una lipasi: L’attività della lipasi è regolata dall’AMP ciclico Glucagone e adrenalina attivano le lipasi Mobilizzazione dei triacilgliceroli Via di ingresso del glicerolo nella glicolisi Prima di essere ossidati gli acidi grassi vengono legati al coenzima A L’adenosina trifosfato (ATP) favorisce la formazione di un legame tioestere tra il gruppo carbossilico di un acido grasso e il gruppo sulfidrilico del CoA. La reazione avviene in due tappe: L’attivazione di un acido grasso avviene in due tappe Questa reazione di attivazione avviene sulla membrana mitocondriale esterna ed è catalizzata dall’enzima acilCoA sintetasi. Acido grasso +CoA + ATP + H2O Æ AcilCoA + AMP + 2 Pi ΔG0’= -32 5 Kj/mole La carnitina trasporta gli acidi grassi attivati nella matrice mitocondriale Enzimi: Carnitina aciltransferasi I e II Ingresso degli acidi grassi nei mitocondri attraverso il trasportatore acil-carnitina/carnitina L’acil carnitina translocasi Fasi dell’ossidazione degli acidi grassi Formazione di un doppio legame Il doppio legame viene idratato per aggiungere un atomo di ossigeno L’alcol viene ossidato a un chetone L’acido grasso viene scisso dal CoA per formare AcetilCoA e una catena di acido grasso di 2 atomi più corta La via di ossidazione degli acidi grassi (β ossidazione) La via di ossidazione degli acidi grassi (β ossidazione) L’equivalente di 2 molecole di ATP viene consumato nell’attivazione del palmitato (ATP scisso a AMP e 2 molecole di Pi), pertanto: L’ossidazione completa di una molecola di palmitato produce 106 molecole di ATP Gli orsi ossidano i loro grassi durante il letargo Con un acido grasso con numero pari di atomi di carbonio, durante l’ultima tappa della β-ossidazione un acetoacetil-SCoA è scisso in 2 Acetil-SCoA O O O CH3 - C - CH2 - C - SCoA + CoASH ¿ 2 CH3 - C- SCoA Acetoacetil-CoA Acetil-CoA Enzima: tiolasi La maggior parte dei lipidi presenti in natura contiene acidi grassi con un numero pari di atomi di carbonio Gli acidi grassi a catena dispari sono presenti in quantità significative nei lipidi delle piante e in alcuni organismi marini Il propionil-SCoA e l’acetil-SCoA sono i prodotti finali della β-ossidazione quando viene ossidato un acido grasso a numero dispari di atomi di carbonio. O O CH3 - CH2 - C - SCoA Propionil-CoA CH3 - CH2 - CH2 - CH2 - C - SCoA O CH3 - C - SCoA Acetil-CoA L’ossidazione degli acidi grassi a catena dispari ha bisogno di altri tre enzimi Struttura del coenzima B12 Formazione di un radicale 5’-deossiadenosilico La reazione della metilmalonil CoA mutasi comincia con la scissione omolitica del legame che unisce lo ione Co3+ a un atomo di C del ribosio della porzione adenosinica. La scissione genera un radicale 5’deossiadenosilico e determina la riduzione del Co3+ a Co2+. Il ruolo del coenzima B12 nelle migrazioni intramolecolari è quello di fungere da sorgente di radiclai liberi per l’estrazione di atomi di idrogeno. Una proprietà essenziale del coenzima è la debolezza del legame cobalto-carbonio, la cui facile scissione genera un radicale Formazione di succinil CoA mediante una reazione di riarrangiamento Il radicale libero generato dal coenzima B12 estrae un atomo di idrogeno nel riarrangiamento del metilmalonil CoA a succinil CoA Ossidazione di un acido grasso monoinsaturo L’ossidazione richiede un altro enzima, l’enoil-CoA isomerasi, che modifica la posizione del doppio legame convertendo l’isomero cis nell’isomero trans. L’intermedio normale della β ossidazione. Ossidazione di un acido grasso poliinsaturo L’ossidazione richiede oltre all’enoil-CoA isomerasi, un secondo enzima ausiliario la 2,4dienol-CoA reduttasi NADPHdipendente. Ossidazione di un acido grasso poliinsaturo (b) L’ossidazione richiede oltre all’enoil-CoA isomerasi,un secondo enzima ausiliario: la 2,4-dienol-CoA reduttasi NADPH-dipendente. L’azione combinata di questi due enzimi converte l’intermedio trans-Δ2,cis-Δ4dienoil-CoA in trans-Δ2enoil-CoA, che può proseguire nella β ossidazione. Per l’ossidazione di qualsiasi acido grasso polinsaturo sono sufficienti 2 enzimi addizzionali I doppi legami in posizione dispari sono convertiti dall’isomerasi; I doppi legami in posizione pari dalla riduttasi e dall’isomerasi. Regolazione allosterica dell’ossidazione degli acidi grassi Il malonil-CoA, il primo intermedio della biosintesi citosolica degli acidi grassi a catena lunga a partire da acetilCoA, inibisce la carnitina aciltransferasi I, impedendo l’accesso degli acil CoA alla matrice mitocondriale nei momenti di abbondanza. Regolando l’acetil CoA carbossilasi, che sintetizza malonil CoA, è possibile regolare in modo coordinato la sintesi e la degradazione degli acidi grassi Il malonil-CoA aumenta quando l’animale è ben rifornito di carboidrati. Regolazione ormonale: Adrenalina e glucagone ½ mobilitazione e rilascio di acidi grassi (+ lipasi) L’insulina si oppone a questo effetto Nelle piante la β ossidazione non è una fonte rilevante di energia metabolica I mitocondri delle cellule delle piante sono privi degli enzimi della β-ossidazione. Durante la germinazione, i triacilgliceroli di riserva sono convertiti in glucosio e in una varietà di metaboliti essenziali. Gli acidi grassi sono ossidati a acetil-CoA nei gliossisomi. L’acetil-CoA viene convertito attraverso il ciclo del gliossilato in precursori a quattro atomi di carbonio che entrano nella gluconeogenesi. Confronto tra la β ossidazione mitocondriale e quella perossisomiale e gliossisomiale Funzione della β ossidazione nella conversione dei triacilgliceroli in glucosio nei semi in germinazione. Acetil - CoA + ossalacetato - ossalacetato (digiuno, diabete: l’ossalacetato viene utilizzato per sintetizzare glucosio) Ciclo dell’acido citrico Formazione di “corpi chetonici” Il destino dell’acetil-CoA dipende dalla disponibilità di ossalacetato per formare citrato (prima reazione del ciclo dell’acido citrico). Corpi chetonici Acetoacetato D-β-idrossibutirrato Formazione dei corpi chetonici dall’acetil-CoA Può avvenire anche non enzimaticamente Il β-idrossibutirrato come fonte di energia L’acetoacetato e il D-β-idrossibutirrato sono trasportati dal sangue ai tessuti extraepatici dove sono ossidati nel ciclo dell’acido citrico. L’acetoacetato e il 3-idrossibutirrato sono combustibili normali della respirazione e sono quantitativamente importanti come fonte energetica Il muscolo cardiaco e la corteccia renale impiegano di preferenza l’acetoacetato anziché il glucosio. Il glucosio, invece, è il principale combustibile del cervello e degli eritrociti negli individui ben nutriti che assumono una dieta bilanciata. Il cervello si adatta all’utilizzo di acetoacetato durante il digiuno e nello stato diabetico L’acetoacetato può essere considerato una forma solubile e trasportabile di unità acetile. Gli acidi grassi vengono rilasciati dal tessuto adiposo e convertiti in unità acetile dal fegato che poi li esporta sotto forma di acetoacetato. Formazione ed esportazione dei corpi chetonici. Le condizioni che determinano un aumento della gluconeogenesi (con sottrazione di ossalacetato dal ciclo dell’acido citrico) esaltano la conversione dell’acetil-CoA in acetoacetato (es.: diabete, digiuno prolungato) Elevate concentrazioni di acetoacetato nel sangue significano un’abbondanza di unità acetile e determinano diminuzione della velocità della lipolisi nel tessuto adiposo. Biosintesi degli acidi grassi Gli acidi grassi sono sintetizzati e degradati attraverso vie differenti La sintesi degli acidi grassi avviene nel citosol; la degradazione nella matrice mitocondriale. Gli intermedi della sintesi sono legati covalentemente ai gruppi sulfidrilici di una proteina trasportatrice di acili (ACP); gli intermedi della degradazione sono legati al coenzima A. La molecola attivata che cede unità bicarboniose nel processo di allungamento è il malonil-ACP. La reazione di allungamento è favorita dal rilascio di CO2. L’agente riducente nella sintesi degli acidi grassi è il NADPH. Negli animali superiori gli enzimi della sintesi degli acidi grassi sono uniti in una unica catena polipeptidica detta acido grasso sintasi. L’allungamento della catena ad opera della acidido grasso sintasi si ferma alla formazione di palmitato (C16). Ulteriori allungamenti e l’inserzione di doppi legami sono effettuati da altri sistemi enzimatici. Il citrato trasporta dai mitocondri al citosol i gruppi acetile che vengono usati per la sintesi degli acidi grassi. Per ogni acetil CoA trasferito dai mitocondri al citosol viene prodotta 1 molecola di NADPH. La formazione di malonil CoA è la tappa di comando nella sintesi degli acidi grassi La biosintesi degli acidi grassi inizia nel citosol con la formazione di malonil CoA catalizzata dalla acetil CoA carbossilasi, che contiene biotina. Come nel caso degli altri enzimi che contengono biotina (es. piruvato carbossilasi) si forma un intermedio carbossibiotina a spese di ATP Il gruppo CO2 attivato viene poi trasferito per formare malonil CoA biotina-enzima + ATP + HCO3- ' CO2-biotina-enzima + ADP + Pi CO2-biotina-enzima + acetil CoA' malonil CoA + biotina-enzima Gli intermedi della sintesi degli acidi grassi sono legati ad una proteina trasportatrice di acili (ACP); il legame avviene al terminale sulfidrilico di una fosfopanteteina, lo stesso gruppo presente nel coenzima A. La proteina che trasporta gli acili Le tappe della degradazione e della sintesi degli acidi grassi La sequenza a quattro tappe che allunga la catena acilica nascente di due atomi di carbonio. La sintesi del palmitato I cicli di allungamento si susseguono fino alla formazione di C16-acil-ACP, che non è più substrato dell’enzima condensante, ma viene idrolizzato a palmitato e ACP. Sequenza delle reazioni che avvengono durante la sintesi di un acido grasso Il complesso dell’acido grasso sintasi è mostrato in modo schematico sotto forma di un disco. Ogni segmento del disco rappresenta una delle sei attività enzimatiche del complesso. Al centro del disco si trova la proteina che trasporta gli acili (ACP), con il braccio fosfopanteteinico. Prima tappa Il gruppo acetilico dell’acetil CoA viene trasferito al gruppo -SH di un residuo di cisteina della β-chetoacil sintasi (KS) Enzima: acetil-CoA-ACP transacetilasi (AT) Seconda tappa Il gruppo malonile viene trasferito dal malonil CoA al gruppo -SH dell’ACP Enzima: malonil-CoA-ACP transferasi (MT) Condensazione Enzima: β-chetoacil-ACP sitasi (KS), o enzima condensante Riduzione Enzima: β-chetoacil-ACP reduttasi (KR) Deidratazione Enzima: β-idrossiacil-ACP (HD) deidratasi Riduzione del doppio legame Enzima: enoil-ACP reduttasi (ER) Traslocazione del gruppo butirrilico sulla Cys della βchetoacil-ACP sintasi (KS) Enzima: Nuovamente l’ acetil-CoA-ACP transacetilasi (AT) che può trasferire gruppi acile diversi dal gruppo acetile Secondo ciclo delle reazioni del complesso dell’acido grasso sintasi. Struttura delle varie acido grasso sintasi L’acido grasso sintasi dei batteri e delle piante è un complesso formato da 7 polipeptidi diversi. Nel lievito le 7 attività sono riunite in 2 polipeptidi. Nei vertebrati tutte le attività sono su di una sola catena polipeptidica L’acido grasso sintasi animale La proteina funzionale è un dimero. Ciascuna delle catene identiche nel dimero contiene tre domini. Il dominio 1 (blu) contiene la acetil transferasi (AT), la malonil transferasi (MT) e l’enzima condensante (CE). Il dominio 2 (giallo) contiene la proteina trasportatrice di acili (ACP), la β-chetoacil riduttasi (KR), la deidratasi (DH) e la enoil reduttasi (ER). Il dominio 3 (rosso) contiene la tioesterasi (TE). Il gruppo flessibile di fosfopanteteina (verde) trasporta la catena di acido grasso da un sito catalitico su una catena all’altro, nonché tra le catene nel dimero. La stechiometria della sintesi del palmitato è: Acetil CoA + 7 malonil CoA + 14 NADPH + 20 H+ Æ palmitato + 7 CO2 + 14 NADP+ + 8 CoA + 6 H2O L’equazione per la sintesi del malonil CoA è: 7 Acetil CoA + 7 CO2 + 7 ATP Æ 7 malonil CoA + 7 ADP + 7 Pi + 14 H+ La stechiometria complessiva per la sintesi di palmitato è: 8 Acetil CoA + 7 ATP + 14 NADPH + 6 H+ Æ palmitato + 14 NADP+ + 8 CoA + 6 H2O + 7 ADP + 7 Pi Il sistema di trasporto dei gruppi acetilici dai mitocondri al citosol, per la sintesi degli acidi grassi L’acetato viene trasportato fuori dai mitocondri sotto forma di citrato Il NADPH è generato dalla via del pentoso fosfato e dall’enzima malico La acetil CoA carbossilasi svolge un ruolo essenziale nella regolazione del metabolismo degli acidi grassi. Acetil CoA Acetil CoA carbossilasi + citrato - palmitoil CoA + insulina - glucagone Malonil CoA Acidi grassi Elevati livelli di citrato indicano che unità a due atomi di carbonio e ATP sono disponibili per la sintesi degli acidi grassi. ½ ATP inibisce isocitrato deidrogenasi = ½ citrato Regolazione della sintesi degli acidi grassi
Scaricare