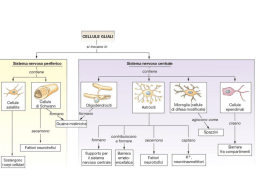
Perché il potenziale di riposo del neurone è –65 mV? La pompa sodio-potassio mantiene in continuazione una differenza di concentrazione di K+ e Na+ tra interno ed esterno della cellula Il sodio, concentrato all’esterno, tende ad entrare mentre il potassio, concentrato all’esterno, tende ad uscire Na+ K+ Nella membrana sono presenti dei canali per il potassio sempre aperti (canali passivi per il K+ ) a riposo (quando praticamente non vi sono altri canali aperti) vi è una uscita lenta ma costante di ioni K+ i quali portano all’esterno cariche positive rendendo negativo l’interno della cellula L’ uscita di ioni K+ si arresta quando le forze elettriche iniziano a risospingere il potassio dentro la cellula (l’esterno è positivo e le cariche positive si respingono). Nella maggior parte dei neuroni questo avviene attorno al valore del potenziale di membrana di circa –65 mV NB: Il potenziale di riposo di una cellula non dipende direttamente dall’azione della pompa sodio-potassio, che si limita a mantenere concentrazioni disuguali di K+ e Na+ ai due lati della membrana. Essa dipende invece dalla differente permeabilità della membrana ai due ioni in questione Esiste un modo preciso per calcolare il potenziale di membrana a riposo (o in qualsiasi altro momento dell’attività del neurone) conoscendo 1) le concentrazioni Interne ed Esterne di ciascun ione (K+, Na+ ecc.) 2) la permeabilità della membrana a ciascun ione in quel momento. Questo modo è mediante l’equazione di Goldman. RT Vm = F ln + [K ] + Pk e PNa[Na ]e + + Pk[K ]i PNa[Na ]i [Cl ] PCl e PCl[Cl ]i Potenziali di membrana • Potenziale di riposo • Potenziali graduati (chiamati anche potenziali locali) • Potenziali post-sinaptici • Potenziali di recettore • Potenziale d’azione (chiamato anche spike o impulso nervoso) Per poter misurare il potenziale di membrana occorre una apparecchiatura costituita da : q Una serie di micro-elettrodi q Un amplificatore del segnale (perché le variazioni sono solo di pochi millivolt) q Uno strumento per visualizzare i potenziali (un voltmetro o, più comunemente, un oscilloscopio) L’oscilloscopio Nell’oscilloscopio le variazioni di voltaggio sono trasformate in movimenti di un pennello di elettroni su uno schermo a fosfori Il movimento di elettroni lascia una traccia sotto forma di un grafico in cui l’asse verticale rappresenta il voltaggio e l’asse orizzontale il tempo che scorre (da sinistra a destra) L’assone gigante del calamaro È utile inoltre utilizzare un neurone di grandi dimensioni. La natura mette a disposizione alcuni neuroni di dimensione giganti. Il modello più utilizzato è il neurone gigante di calamaro il cui assone misura quasi un millimetro di diametro Ogni calamaro ne ha due e gli servono per comandare il sistema di fuga L’assone gigante, isolato dal calamaro può sopravvivere per un giorno o due in soluzione fisiologica Oscilloscopio Elettrodo di riferimento Amplificatore Assone Microelettrodo per la misura del potenziale di membrana I microelettrodi Un microelettrodo è un elettrodo così sottile da riuscire a penetrare la membrana plasmatica senza lesionarla. Non può essere di metallo perché a quelle dimensioni esso risulterebbe troppo fragile Per costruirlo si parte invece da un tubicino di vetro di pochi mm di diametro. Questo viene scaldato finché diviene malleabile e quindi viene ‘tirato’ fino a divenire estremamente sottile (ma per quanto sottile rimane cavo al suo interno) Il vetro non conduce la corrente, tuttavia se si riempie il suo interno di una soluzione salina concentrata esso diventa un conduttore di corrente sufficientemente buono da misurare i potenziali di membrana di una cellula Il potenziale di riposo 0 Se entrambi gli elettrodi sono posti nel liquido extracellulare non si registra differenza di potenziale -65 Quando il microelettrodo viene introdotto nel neurone del calamaro l’oscilloscopio registra un potenziale attorno a –65 mV Il potenziale di riposo In tutte le cellule del corpo l’interno è sempre più negativo dell’esterno A riposo il potenziale di membrana di una cellula è normalmente compreso tra i –40 mV e –90 mV I potenziali graduati In condizioni normali sono generati in due modi A livello dei recettori dall’azione di stimoli fisici (suoni, luce, pressione) Potenziali di recettore A livello delle sinapsi dall’azione dei neuroni presinaptici Potenziali post-sinaptici eccitatori (EPSP) Potenziali post-sinaptici inibitori (IPSP) Sperimentalmente i potenziali graduati possono essere simulati mediante una debole corrente trasmessa con un elettrodo Per misurare i potenziali graduati occorre munirsi di un secondo microelettrodo collegato con un generatore di stimoli capace di produrre corrente di debole intensità (elettrodo stimolatore) La situazione ideale per studiare le proprietà dei potenziali graduati è quella di posizionarsi a livello di un dendrite (non c’è mielina e non si manifestano altri tipi di potenziale) Quattro proprietà dei potenziali graduati +10 0 Corrente applicata -10 -60 -65 0 Potenziale di membrana -70 1) La variazione del potenziale è proporzionale all’intensità della corrente applicata (indipendentemente dalla polarità) 2) Dopo la cessazione dello stimolo il potenziale ritorna gradualmente al valore di riposo Elettrodo stimolatore T1 Dendrite T1 T1 3) Il potenziale viene registrato in maniera pressoché simultanea in ogni punto del dendrite T1 4) L’intensità del potenziale decresce man mano che ci allontaniamo dal punto di stimolazione Quando la cellula passa da un valore negativo (il potenziale di riposo) ad uno ancor più negativo si dice che si iperpolarizza Quando la cellula passa da un valore negativo (il potenziale di riposo) ad uno meno negativo si dice che si depolarizza Il potenziale d’azione -65 Iperpolarizzazione Se si somministrano stimoli iperpolarizzanti (cioè che rendono la cellula ancor più negativa) via via maggiori la cellula risponde in modo proporzionalmente maggiore -65 Depolarizzazione Se si somministrano stimoli depolarizzanti (cioè che rendono la cellula meno negativa) si osserva un comportamento speculare Quanto tuttavia si supera un valore di soglia di circa –50 o –55 mV, si assiste ad un fenomeno nuovo, il potenziale d’azione i potenziali graduati avvengono a livello dei dendriti e del soma per poi essere integrati nella zona trigger del neurone dove troviamo canali voltaggio dipendenti per Na+ (assenti su dentriti) + 40 Il potenziale d’azione 0 Soglia -55 -65 Potenziale di riposo -100 2msec Nello spazio di meno di due millisecondi l’interno della cellula diviene positivo (fino a circa + 40 mV) e poi ritorna ad un valore negativo, prossimo a quello del potenziale di riposo Propagazione del potenziale d’azione Disponendo più elettrodi lungo l’assone è possibile studiare in che modo il potenziale d’azione si propaga lungo l’assone 1 Durante la propagazione, del potenziale d’azione non c’è attenuazione del segnale (come invece avviene per i potenziali graduati) Si tratta di un segnale stereotipato 2 3 1 2 3 Il potenziale d’azione non è simultaneo in tutta la cellula ma si propaga con una velocità tra 1 e 100 metri al secondo (a seconda delle caratteristiche del neurone) Cosa succede se si stimola un assone più intensamente? Stimolazione debole Se si aumenta l’intensità della stimolazione, il potenziale d’azione compare sempre con intensità caratteristica (ad esempio sempre +40mV) Tuttavia aumentando l’intensità della stimolazione, aumenta la frequenza con cui i potenziale d’azione si susseguono Stimolazione intensa Nel sistema nervoso la codifica della intensità di un segnale non è data dall’ampiezza del potenziale d’azione ma dalla sua frequenza In sintesi: I potenziali graduati si propagano pressochè istantaneamente ma decrescono di intensità con la distanza Il potenziale d’azione si propaga con una velocità misurabile ma l’ampiezza della variazione di potenziale rimane costante indipendentemente dalla distanza e dalla intensità dello stimolo che lo ha provocato
Scaricare