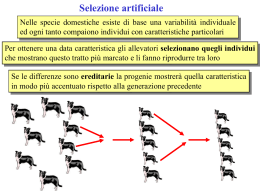
La genetica di popolazione Il caso e la necessità Prima parte Liceo Scientifico “A. Meucci” di Aprilia Prof. Neri Rolando Da Darwin alla sintesi moderna Le popolazioni sono le unità su cui agisce l’evoluzione – – La popolazione (un gruppo di individui della stessa specie che vivono nello stesso posto nello stesso momento) rappresenta l’insieme più piccolo di organismi soggetto all’evoluzione. Una specie è un gruppo di individui, generalmente concentrati in popolazioni, che sono in grado di incrociarsi tra loro e produrre prole fertile. Genetica delle popolazioni – La genetica delle popolazioni, nata intorno al 1920, è la scienza che si occupa dei cambiamenti genetici delle popolazioni. – La sintesi moderna (o teoria sintetica dell’evoluzione), sviluppatasi all’inizio degli anni Quaranta, è una teoria evolutiva che considera le popolazioni come le unità dell’evoluzione e tiene conto di gran parte dei concetti espressi da Darwin. Il pool genico e la microevoluzione – L’insieme di tutti gli alleli di tutti gli individui che compongono una popolazione, presenti in qualsiasi momento, costituisce il pool genico della popolazione. – La microevoluzione è un cambiamento nella frequenza relativa degli alleli nel pool genico di una popolazione. Le basi della teoria sintetica -Tutti gli organismi discendono da un unico capostipite - Nascono più individui di quanti ne possano sopravvivere - La variabilità individuale è frutto delle mutazioni che, attraverso ricombinazioni alleliche, interazioni geniche e crossing-over (meiosi e fecondazione), arricchiscono le diverse forme che ogni carattere può assumere - L’evoluzione è un fenomeno di popolazione e non opera su un genotipo ma sull’intero patrimonio genetico (pool genico) attraverso i fenotipi - La selezione naturale preserva le mutazioni vantaggiose, i cui portatori aumenteranno di frequenza da una generazione all’altra, ed elimina più o meno rapidamente quelle svantaggiose. Non evoluzione In una popolazione che non si evolve il pool genico rimane immutato nel corso delle generazioni In una popolazione che non si evolve il mescolamento di geni che accompagna la riproduzione sessuata non altera la composizione genetica della popolazione. Zampa con membrana Zampa senza membrana Le basi genetiche dell’evoluzione Nonostante gli studi di Mendel non fossero ancora noti, Darwin aveva intuito che i caratteri di un individuo erano trasmissibili alla generazione successiva, ma non era riuscito a capire quale fosse il meccanismo secondo il quale l’informazione genetica veniva trasmessa di generazione in generazione. Questa trasmissibilità dei caratteri, tuttavia, è un punto fondamentale del meccanismo della selezione naturale come spiegazione della teoria dell' evoluzione. Ma, in realtà, c’era anche un altro punto focale della sua Teoria che aveva lasciato in sospeso: quale o quali fossero le cause della incredibile variabilità che presentano le specie. LINK: la mappa della Teoria dell’evoluzione di Darwin Diversità genetica La variabilità della struttura genetica delle popolazioni viene solitamente misurata analizzando i polimorfismi della sequenza di DNA e individuando i diversi genotipi. La genetica di popolazione studia la composizione dei geni all’interno di un gruppo di individui cioè le frequenze alleliche e i diversi genotipi, le variazioni che interessano questi geni nel tempo e le differenze esistenti tra vari gruppi o popolazioni Le basi genetiche dell’evoluzione La teoria dell’evoluzione di Darwin, quindi, aveva un grande difetto: non spiegava l’origine della variazione e come particolari varianti venissero ereditate. Nel 1900, la riscoperta dei principi di Mendel fornì una spiegazione a posteriori: i caratteri sono determinati dai geni, che segregano alleli differenti, e i geni – cioè le informazioni – sono trasmessi alla progenie attraverso i gameti prodotti dai genitori. Lo studio della trasmissione genetica in incroci sperimentali e negli alberi genealogici diede origine alla genetica delle popolazioni (Wright S., Fisher R.A. e Halden B.S., 1930) che divenne il fondamento della teoria evolutiva di Darwin, con il termine di «Teoria Sintetica dell’Evoluzione» . Le basi genetiche dell’evoluzione La struttura genetica delle popolazioni cambia nel tempo attraverso le generazioni. Le cause di tali cambiamenti sono: • la selezione naturale o la deriva genetica; • la selezione artificiale. I parametri per descrivere la struttura genetica di una popolazione sono: • le frequenze alleliche; • le frequenze genotipiche. La formula dell’evoluzione Nel suo libro «Perché io credo in Colui che ha fatto il mondo» (Il saggiatore, Milano, 1999), il fisico Antonino Zichichi dismette la teoria biologica dell’evoluzione come non scientifica, in quanto non espressa da un’equazione. Tra le tante critiche che le sono state rivolte nel secolo e mezzo che ci separa dalla pubblicazione di «L’origine delle specie» di Charles Darwin, questa è una delle più disinformate: la matematica dell’evoluzionismo esiste infatti da un secolo esatto, visto che fu trovata indipendentemente nel 1908 dal matematico inglese Godfrey Hardy e dal medico tedesco Wilhelm Weinberg, da cui il nome di legge di Hardy-Weinberg con cui è (o dovrebbe essere) conosciuta. La formula dell’evoluzione Le leggi dell’ereditarietà erano state scoperte nel 1866 da Gregor Mendel, e riscoperte nel 1900 da Hugo de Vries, Karl Korrens ed Erik von Tschermak: sostanzialmente stabiliscono che se si incrociano due individui eterozigoti, portatori cioè di due varianti A e a di uno stesso carattere (chiamate tecnicamente alleli), il primo dominante e il secondo recessivo, allora la loro prole sarà per ¼ omozigote dominante (AA), per ¼ omozigote recessiva (aa), e per metà eterozigote (aA o Aa), semplicemente per le leggi della combinatoria. Infatti la meiosi separa i due alleli di ciascun genitore, così che metà dei gameti (gli spermatozoi maschili e gli ovuli femminili) ne avranno uno, e metà l’altro. E, poiché la fecondazione combina casualmente un gamete maschile e uno femminile, ciascuna delle quattro combinazioni avrà ¼ delle possibilità. LINK: Le leggi di Mendel La formula dell’evoluzione Nel 1902 Udny Yule sollevò una possibile obiezione nei confronti del mendelismo: se la prole esprime per ¾ la variante dominante del carattere (che richiede solo un allele A), e per ¼ quella recessiva (che richiede entrambi gli alleli a), come mai gli alleli e i caratteri dominanti non aumentano nella popolazione, fino a far scomparire quelli recessivi? La formula dell’evoluzione La risposta fu data dal matematico inglese Godfrey Hardy in una lettera a «Science» intitolata «Proporzioni mendeliane in una popolazione mista», che iniziava modestamente così: «Sono riluttante a intromettermi in una discussione su un argomento di cui non sono esperto, e mi sarei aspettato che il semplice fatto che voglio far notare fosse familiare ai biologi. Ma alcune osservazioni di Udny Yule suggeriscono che possa valer la pena farlo». La formula dell’evoluzione Equilibrio di Hardy-Weinberg Il semplice fatto era che se i due alleli A e a sono rispettivamente presenti nei gameti della popolazione nelle percentuali p e q, allora lo stesso ragionamento di prima mostra che se gli individui si accoppiano in maniera casuale la loro prole sarà omozigote dominante in proporzione p2, omozigote recessiva in proporzione q2 ed eterozigote in proporzione 2pq, cioè: P2 + 2pq + q2 = 1 Ma allora nei gameti della prole l’allele A sarà presente in proporzione p2 + pq = p • (p + q), e l’allele a in proporzione q2 + pq = q • (p + q). E poiché p + q = 1, le due percentuali sono le stesse di quelle di partenza, cioè p e q. In altre parole le percentuali sia dei due alleli sia dei loro genotipi (cioè, degli individui che esibiscono la rispettiva variante del carattere), rimangono costanti di generazione in generazione, indipendentemente dai valori di partenza. La formula dell’evoluzione Equilibrio di Hardy-Weinberg In sintesi: Dall’incrocio tra eterozigoti ( Aa x Aa ) si ha la seguente somma di genotipi AA + 2 Aa + aa Se p = frequenza dell’allele A e q = frequenza dell’allele a Allora diventa pp + 2 pq + qq Cioè p2 + 2 pq + q2 Quindi l’allele A è presente nella proporzione p2 + pq = p (p + q) E l’allele a in proporzione q2 + 2 pq = q (p + q) E poiché p+q=1 Allora le due percentuali sono le stesse di quelle di partenza, cioè p e q. Nella pagina successiva viene mostrato un esempio concreto di ciò. La formula dell’evoluzione Per verificare l’equilibrio di Hardy-Weinberg si possono seguire gli alleli in una popolazione. La farfalla notturna Biston betularia. si presenta in due forme alternative determinate da un unico gene. L'allele A determina il colore scuro e ha una dominanza completa sull'allele a che codifica per il colore chiaro. Supponiamo che analizzando una popolazione di 500 individui si trovi la seguente composizione. E’ possibile ricavare le frequenze dei tre genotipi AA, Aa, e aa. E’ possibile inoltre calcolare le frequenze alleliche p e q. Nel momento della meiosi le farfalle producono dei gameti contenenti gli alleli A e a con queste stesse frequenze. Nel momento della fecondazione la combinazione tra i gameti avviene in modo casuale secondo la regola del quadrato di Punnett. L'equilibrio di Hardy-Weinberg è rispettato in quanto le frequenze genotipiche della prima generazione sono uguali a quelle della popolazione originaria. La formula dell’evoluzione Equilibrio di Hardy-Weinberg Fenotipi Definizione: L’ equilibrio di HardyWeinberg stabilisce che il mescolamento dei geni durante la riproduzione sessuata non altera le frequenze dei diversi alleli in un pool genico. Genotipi Numero di animali (totale 500) Frequenze genotipiche Numero di alleli del pool genetico (totale 1000) Frequenze alleliche WW 320 ww 160 20 160 0,32 500 320 0,64 500 640 W Ww 160 W 160 w 800 0,8 W 1000 20 0,04 500 40 w 200 0,2 w 1000 La formula dell’evoluzione Equilibrio di Hardy-Weinberg Come già mostrato in precedenza per verificare l’equilibrio di HardyWeinberg si possono seguire gli alleli in una popolazione. Ricombinazione degli alleli della prima generazione (genitori) Gameti maschili W maschile w maschile p 0,8 q 0,2 W femminile p 0,8 Gameti femminili p2 WW 0,64 wW w femminile q 0,2 qp 0,16 Ww pq 0,16 q2 ww 0,04 Seconda generazione: Frequenze genotipiche Frequenze alleliche 0,64 WW 0,32 Ww 0,8 W 0,04 ww 0,2 w La formula dell’evoluzione Equilibrio di Hardy-Weinberg L’equazione di Hardy-Weinberg è utile nello studio delle malattie genetiche – I consultori genetici utilizzano l’equazione di Hardy-Weinberg per stimare la percentuale dei soggetti portatori di alleli responsabili di alcune malattie ereditarie. – Conoscere la frequenza di un allele dannoso è utile per qualunque programma di sanità pubblica che si occupi di malattie genetiche. Le forze evolutive Poiché l’evoluzione avviene quando le cose cambiano, essa è possibile soltanto quando non si verificano almeno alcune delle cinque condizioni che portano all’equilibrio di Hardy e Weinberg. Tre di queste condizioni sono di natura statistica: • gli accoppiamenti devono essere equiprobabili • e casuali, • e la popolazione deve essere sufficientemente grande perché valga la legge dei grandi numeri. Le altre due condizioni sono di natura fisica: • il sistema deve essere stabile • e chiuso. Le forze evolutive • Ora, accoppiamenti non equiprobabili sono tipici della “selezione naturale”, che favorisce la fecondità o la sopravvivenza di alcuni individui rispetto ad altri. • Accoppiamenti non casuali sono invece tipici della “selezione artificiale” imposta da allevatori o coltivatori, così come della “selezione sessuale” praticata da partner selettivi. • Se la popolazione è piccola si può verificare una “deriva genetica”, per esempio nei pochi sopravvissuti a un cataclisma (“collo di bottiglia”), o in pochi emigrati distaccatesi da un gruppo (“effetto del fondatore”). • Instabilità del sistema, infine, possono essere provocate da mutazioni endogene (copiatura) o esogene (danneggiamento) dei singoli alleli, • e aperture del sistema possono essere provocte da flussi genici in entrata nella popolazione (immigrazione) o in uscita (emigrazione). Le forze evolutive Deriva genetica Selezione Migrazione Accoppiamento non casuale Mutazione La mutazione crea variabilità genetica, le altre forze evolutive agiscono sulla variabilità creata dalla mutazione ed eventualmente dalla ricombinazione genetica dovuta alla riproduzione sessuale (crossing-over, assortimento indipendente dei cromosomi, fecondazione). Le forze evolutive 1. 2. 3. 4. 5. Deriva genetica Selezione Migrazione Mutazione Accoppiamento non casuale (inincrocio ed accoppiamento assortativo) Le forze evolutive 1-4 agiscono causando una variazione delle frequenze alleliche e genotipiche nelle popolazioni. L’accoppiamento non casuale determina una variazione delle sole frequenze genotipiche rispetto all’equilibrio di Hardy-Weinberg, perché agiscono direttamente su determinati fenotipi. Quali sono gli effetti delle forze evolutive sul grado di variabilità intra- ed interpopolazioni? Le forze evolutive Le mutazioni generano nuovi alleli e la riproduzione sessuata li ricombina L’origine della variabilità genetica è la mutazione. Una mutazione è qualsiasi cambiamento nel DNA di un organismo. Le mutazioni naturali possono avere cause diverse, ma hanno una caratteristica in comune: sono casuali rispetto ai bisogni adattativi dell’organismo. In genere le mutazioni sono dannose o ininfluenti per chi le porta ma, se le condizioni ambientali cambiano, i nuovi alleli possono rivelarsi vantaggiosi. In natura le mutazioni si verificano con una frequenza molto bassa. continua Le forze evolutive Le mutazioni generano nuovi alleli e la riproduzione sessuata li ricombina Se è vero che le mutazioni introducono nuovi alleli in una popolazione, è difficile che possano consentirne la diffusione. Almeno per gli eucarioti, questo è piuttosto l’esito dei fenomeni legati alla meiosi e alla riproduzione sessuata. Questi due ultimi processi non generano di per sé nuovi alleli, ma favoriscono la ricombinazione (▶figura pagina successiva), vale a dire la formazione di nuove associazioni tra gli alleli esistenti. Per quanto possa sembrare sorprendente, questo «rimescolare le carte» è spesso più proficuo, in termini evolutivi, di quanto non sia la comparsa di nuovi alleli. Insomma, generalmente l’esito positivo o negativo di una partita dipende dalla combinazione di carte che un giocatore possiede. continua Le forze evolutive Il ruolo delle mutazioni – Anche le mutazioni (cambiamenti casuali nel DNA di un organismo che possono dare origine a un nuovo allele) possono determinare microevoluzione. – Le mutazioni sono la causa principale della variabilità genetica e rappresentano il punto di partenza dei processi evolutivi. Le forze evolutive Il flusso genico può cambiare le frequenze alleliche È raro che una popolazione sia completamente isolata da altre popolazioni della stessa specie. Più spesso si assiste alla migrazione di individui o allo spostamento di gameti da una popolazione all’altra, due fenomeni che insieme costituiscono il flusso genico. Gli individui o i gameti introdottisi nel nuovo ambiente possono apportare al pool genico della popolazione alleli nuovi. Anche se questo non accade, se le due popolazioni hanno frequenze alleliche diverse il flusso può comportare un cambiamento delle frequenze degli alleli nella popolazione originaria. continua Le forze evolutive Il flusso genico è un altro fattore che può determinare microevoluzione: • si verifica quando individui fertili entrano a fare parte di una popolazione o se ne allontanano, oppure quando si verifica un trasferimento di geni; • tende a ridurre le differenze genetiche tra le popolazioni. continua Le forze evolutive Il flusso genico può cambiare le frequenze alleliche Questo fenomeno cambia le frequenze alleliche nelle popolazioni interessate, mentre le frequenze geniche nell’intera specie non cambiano. Gli Stati Uniti sono un esempio di notevole flusso genico, come anche la Sicilia: sono zone geografiche che hanno ospitato nel tempo individui provenienti da molte diverse popolazioni, in origine lontane tra loro. Si stima che il flusso genico dai bianchi-caucasici americani ai neri americani si è verificato a una media del 3,6% per generazione. Considerando che l'inizio della migrazione dall'Africa all'America è stato circa 300 anni fa, ossia 10 generazioni, si ricava che circa il 30% dei geni degli afroamericani è derivato da caucasici. Le forze evolutive Nelle popolazioni poco numerose la deriva genetica può produrre grandi cambiamenti La deriva genetica agisce nelle popolazioni naturali (di dimensione finita) e consiste nella fluttuazione casuale delle frequenze alleliche di generazione in generazione La variazione delle frequenze alleliche è tanto maggiore quanto più piccola è la popolazione Con il passare del tempo, un allele ha due sole possibilità: estinguersi o fissarsi (si estingue l’altro allele) La probabilità che ha un allele di andare incontro a fissazione è pari alla sua frequenza iniziale Il tempo medio di fissazione di un allele è proporzionale alla dimensione della popolazione continua N = 1000 N = 1000 N = 10000 p = 0.2 N = 100 La deriva genetica La deriva riduce la variabilità entro popolazioni e aumenta quella fra popolazioni Simulazione di deriva genetica in popolazioni diploidi di 10000 individui e di 4 individui continua Le forze evolutive La deriva genetica Consideriamo, ad es., un allele a, che mostra una frequenza dell’1%. In una popolazione di un milione di individui saranno presenti nel pool genico 20000 alleli a. (Ogni individuo diploide porta 2 alleli per un dato gene: nel pool genico di questa popolazione ci sono 2 milioni di alleli per questo particolare gene, e di essi l’1% è costituito dall’allele a. Se per caso alcuni individui di questa popolazione morissero prima di lasciare discendenti, l’effetto di tale evento sulla frequenza dell’allele a sarebbe trascurabile. Invece, in una popolazione di 50 individui la situazione sarebbe diversa in quanto, in questa popolazione, sarebbe presente solo una copia dell’allele a. Se l’unico individuo che porta questo allele non riuscisse a riprodursi o se morisse prima di aver generato una prole, l’allele a andrebbe perso del tutto. Analogamente, se fossero eliminati 10 dei 49 individui prive dell’allele a, la frequenza di a passerebbe da 1/100 a 1/80 continua Le forze evolutive La deriva genetica Per capire questa situazione è necessario ricordare che, quando si studia un fenomeno, la frequenza osservata F0 ( o Legge dei grandi numeri) è uguale al rapporto tra il numero degli eventi favorevoli f e il numero totale dei casi osservati n (F0 = f / n), mentre la probabilità teorica PT è uguale al rapporto tra il numero degli eventi favorevoli f e il numero degli eventi tutti ugualmente possibili n ( PT = f / n). Ad es., immaginiamo di avere un sacchetto contenente 1000 palline di 4 colori diversi (rosso, bianco, giallo, nero) e distribuite in 4 gruppi di 250 palline ciascuno. La PT di estrarre una pallina rossa sarà PT = ¼ = 25%. Tuttavia, se facciamo solo 4 estrazioni, è difficile che la FO sia uguale a PT, infatti, potrei anche non estrarre alcuna pallina rossa; se faccio 40 estrazioni, è probabile che il valore della FO si avvicini a quello della PT. Ma, se le estrazioni sono 400 è quasi certo che i due valori coincidano. Analogamente, è molto probabile che la composizione del pool genico di una popolazione sottoposta a deriva genetica, cioè ad una drastica diminuzione casuale dei suoi membri, sia molto diversa da quella del pool genico della più ampia popolazione originaria. continua Le forze evolutive La deriva genetica Come esempio, supponiamo di far accoppiare dei moscerini della frutta eterozigoti. L’incrocio Aa x Aa produce una generazione F1 nella quale p = q = 0,5 e le frequenze genotipiche sono 0,25 per AA, 0,50 per Aa e 0,25 per aa. Se al momento di produrre la generazione F2 scegliamo a caso 4 individui (che corrispondono a 8 copie del gene) della generazione F1, può capitare che in questa piccola popolazione campione le frequenze alleliche siano diverse da p = q = 0,5. Per esempio, se casualmente abbiamo estratto 2 omozigoti AA e 2 eterozigoti (Aa), le frequenze alleliche del campione saranno p = 0,75 (6 su 8) e q = 0,25 (2 su 8). Ripetendo 1000 volte l’esperimento di campionamento, si otterranno 1000 popolazioni campione delle quali circa 8 risulteranno completamente prive di uno dei due alleli. continua Le forze evolutive Perché è importante la deriva genetica? Importanza evolutiva: cambiamento non adattativo, specie in piccole popolazioni Importanza per la conservazione: perdita di diversità genetica, specie in piccole popolazioni Importanza biomedica: alleli patologici, altrove rari, possono essere comuni in piccole popolazioni continua Le forze evolutive Consideriamo ora due esempi di deriva genetica: l’effetto collo di bottiglia e l’effetto del fondatore Collo di bottiglia Effetto del fondatore Colori diversi indicano alleli diversi ad un ipotetico locus multiallelico continua La deriva genetica L’effetto collo di bottiglia si verifica quando una popolazione subisce ingenti perdite È possibile che popolazioni solitamente numerose di tanto in tanto attraversino periodi difficili, nei quali sopravvive soltanto un piccolo numero di individui. Durante queste fasi di contrazione numerica della popolazione (a causa di malattie, per la caccia ad opera dell’uomo, in seguito a periodi di siccità, ecc…), note come colli di bottiglia, la deriva genetica può portare a una riduzione della variabilità genetica. Il meccanismo è illustrato nella ▶figura A (pagina successiva), nella quale i fagioli rossi e gialli rappresentano i due diversi alleli di un gene. Nel piccolo campione prelevato dalla popolazione iniziale, per puro caso la maggior parte dei fagioli è rossa, così nella popolazione «sopravvissuta» la frequenza dei fagioli rossi è molto più alta che nella popolazione originale. Parlando di una popolazione reale, si potrebbe dire che le frequenze alleliche sono andate alla «deriva». È probabile che una popolazione costretta a passare attraverso un collo di bottiglia perda gran parte della propria variabilità genetica. Un esempio di collo di bottiglia è offerto dai ghepardi (Acinonyx jubatus; (▶figura B). Durante l’ultima era glaciale, questa specie arrivò molto vicina all’estinzione e ne sopravvissero probabilmente poche unità. Questo ha fatto sì che la varietà genetica tra i ghepardi sia molto bassa; ciò è confermato dal fatto che è possibile eseguire un trapianto di pelle tra due ghepardi senza che vi sia alcun rigetto, tanto il patrimonio genetico di donatore e ricevente sono simili. continua Effetto collo di bottiglia continua Deriva genetica Effetto collo di bottiglia Di regola, come è già stato sottolineato, l’effetto «collo di bottiglia» riduce la variabilità genetica complessiva di una popolazione. Un esempio di questo concetto è la potenziale perdita di variabilità tra individui, e dunque della adattabilità, nelle popolazioni di specie a rischio di estinzione che hanno subito l’effetto «collo di bottiglia». Il Ghepardo subì un secondo effetto «collo di bottiglia» nel diciannovesimo secolo quando l’uomo gli ha dato la caccia fin quasi all’estinzione. Attualmente soltanto tre piccole popolazioni sopravvivono in natura, la variabilità genetica è molto bassa rispetto a quella di altri mammiferi, perché per generazioni e generazioni si sono incrociati tra di loro. Anche il Camoscio d’Abruzzo ha subito lo stesso iter ...…. Ma anche per l’uomo si hanno degli esempi. Fra gli Ebrei Ashkenazim l’elevata frequenza della malattia di Tay-Sachs (causa di degenerazione del sistema nervoso nell’infanzia) è attribuita al fatto che la popolazione durante il Medio Evo venne perseguitata e sterminata provocando il fenomeno del collo di bottiglia. Molto probabilmente l’allele recessivo per questa malattia era presente, casualmente, con una frequenza superiore a quella normalmente osservata nella popolazione ebraica, proprio a causa del genocidio subito. continua La deriva genetica L’effetto del fondatore si verifica quando pochi individui pionieri colonizzano una nuova regione Quando alcuni individui colonizzano un nuovo ambiente, è improbabile che portino con sé tutti gli alleli presenti nella popolazione di origine. Il cambiamento che si verifica nella variabilità genetica prende il nome di effetto del fondatore ed è equivalente a quanto accade in una grande popolazione decimata da un collo di bottiglia. continua La deriva genetica In realtà c’è un altro esempio di deriva genetica, dovuto esclusivamente all’azione dell’uomo, ed è la costituzione dei parchi naturali. Queste aree protette rappresentano, è vero, un sistema di conservazione degli habitat naturali, tuttavia, sono anche il risultato della drastica riduzione degli habitat naturali ed abitate da popolazioni piccole, caratterizzate, generalmente, da una limitata varietà genetica. continua Quand’è che una popolazione può dirsi variabile? A N alleli = 5 B N alleli = 2 Quando il genotipo individuale è difficile da prevedere continua Flusso genico e deriva hanno effetti opposti Il flusso genico – introduce nuovi alleli nelle sottopopolazioni e – riduce le differenze fra sottopopolazioni Flusso genico deriva Le forze evolutive L’accoppiamento non casuale modifica le frequenze genotipiche Abbiamo assunto, per la dimostrazione della legge di Hardy-Weinberg, che gli individui si accoppiano a caso e che la scelta del partner non è influenzata dagli alleli che possiede. Tuttavia, se determinati alleli sono responsabili di caratteristiche fenotipiche che influenzano la scelta del partner, questa non avviene in modo casuale, e il fenomeno, detto accoppiamento assortativo, cambia le frequenze genotipiche osservate rispetto a quelle attese dal modello della «popolazione mendeliana». continua Le forze evolutive L’accoppiamento non casuale modifica le frequenze genotipiche Un caso particolare di accoppiamento assortativo è l'inincrocio, fenomeno per cui l’accoppiamento tra parenti avviene con una frequenza più elevata di quella dovuta al caso (ad es., l’autoimpollinazione nelle piante). Negli animali l’accoppiamento non casuale è spesso guidato dal comportamento. Ad es., l’oca selvatica può essere sia bianca sia azzurra; le oche bianche tendono ad accoppiarsi con altre oche bianche e lo stesso fanno le azzurre. Perciò, supponendo che siano interessati solo due alleli, ci sarà una diminuzione nella frequenza degli eterozigoti (rappresentati da 2pq nell’equazione H-W) e un conseguente aumento delle frequenze dei due omozigoti (p2 e q2). È bene sottolineare che l’accoppiamento non casuale altera le frequenze genotipiche, ma non le frequenze alleliche, e quindi non produce adattamento. Esiste però anche una forma particolarmente importante di accoppiamento non casuale, capace di cambiare le frequenze alleliche: la selezione sessuale. continua Le forze evolutive La selezione sessuale influenza il successo riproduttivo La selezione sessuale è un tipo particolare di selezione naturale che agisce sulle caratteristiche che determinano il successo riproduttivo. Consiste nella lotta tra membri di un sesso (di solito maschile) per la conquista dell’atro sesso. Questo tipo di selezione genera il dimorfismo sessuale fenomeno che riguarda le differenze tra maschi e femmine nella modalità di procurarsi un compagno (diversa modalità di «attirare l’attenzione»). Si distingue tra selezione intrasessuale e intersessuale, la prima è una competizione diretta tra gli individui dello stesso sesso (nei vertebrati generalmente i maschi con i rituali di corteggiamento); la seconda è nota anche come scelta del compagno riproduttivo, ovvero diventa prioritario quello di risultare più attraente agli occhi dei membri dell’altro sesso (è il caso, ad es., del pavone). Questo secondo tipo risulta veramente interessante, perché generalmente si pensa che solo l’uomo abbia la capacità di operare scelte. continua Le forze evolutive La selezione sessuale influenza il successo riproduttivo La selezione sessuale, quindi, riguarda soltanto il successo riproduttivo. Ovviamente un animale per arrivare a riprodursi deve sopravvivere, ma se sopravvive e non si riproduce, non dà alcun contributo alla generazione successiva. Non è dunque impossibile che la selezione sessuale favorisca alcune caratteristiche capaci di accrescere la capacità riproduttiva del portatore, anche se ne riducono la capacità di sopravvivenza. Intuitivamente però questo concetto è difficile da comprendere e accettare, ed è stato ignorato o criticato per molti decenni, finché recentemente alcune ricerche sperimentali non ne hanno dimostrato l’importanza. Un esempio è l’enorme coda del maschio dell’uccello vedova (Euplectes progne), che vive in Africa e ha una coda più lunga della testa e del corpo messi insieme. continua Le forze evolutive La selezione sessuale influenza il successo riproduttivo Normalmente i maschi di uccello vedova scelgono un territorio dove eseguire i rituali di corteggiamento per attrarre le femmine, e lo difendono dagli altri maschi. Tutti i maschi, tanto quelli a coda lunga quanto quelli a coda corta, erano capaci di difendere il loro territorio di corteggiamento, a dimostrazione che la lunghezza della coda non incide sulla competizione tra maschi. Tuttavia, i maschi con la coda allungata artificialmente attraevano circa il quadruplo delle femmine dei maschi con la coda accorciata (▶figura nella pagina successiva). continua La selezione sessuale La selezione sessuale influenza il successo riproduttivo Le femmine di uccello vedova preferiscono i maschi con la coda lunga perché se possono sviluppare e conservare una caratteristica così «costosa», nonostante la riduzione di capacità di volo che essa provoca, devono essere per forza sani e vigorosi. Una caratteristica costosa fornisce al sesso che opera la scelta del partner (di solito la femmina) un indizio attendibile per riconoscere gli individui realmente dotati di buone qualità riproduttive da quelli che bluffano. Se le femmine scegliessero il compagno sulla base di una caratteristica facilmente simulabile, non ne ricaverebbero alcun beneficio in termini di fitness.
Scaricare