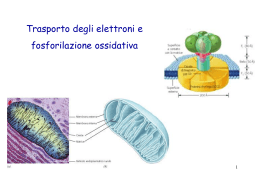
Trasporto degli elettroni e fosforilazione ossidativa 1 2 Gli organismi aerobici durante il processo di ossidazione dei carburanti metabolici consumano ossigeno e producono biossido di carbonio: C6H12O6 + 6O2 6CO2 + 6 H2O suddivisa in due semireazioni: C6H12O6 + 6O2 6CO2 + 12 H+ + 12 eOssidazione degli atomi di C del glucosio 6O2 + 12 H+ + 12 e- 6 H2O Riduzione dell’ossigeno molecolare 3 Gli e- rilasciati durante l’ossidazione del glucosio non sono trasferite direttamente all’O2 ma ai coenzimi NAD+ e FAD tramite i coenzimi, gli e- passano alla catena di trasporto degli e- 4 Durante il processo di trasporto degli elettroni accadono i eguenti eventi: 1) NADH e FADH2 sono riossidati a NAD+ e FAD 2) il trasferimento degli e- partecipa all’ossidazione-riduzione di 10 centri redox localizzati in 4 complessi enzimatici 3) durante il trasferimento degli e-, dal mitocondrio vengono espulsi dei protoni con la generazione di un gradiente protonico attraverso la membrana mitocondriale. 5 6 Struttura di un mitocondrio 80% proteine 50% lipidi 50% proteine 7 NAD+, NADH, NADP+, NADPH, FAD, FADH2 non possono attraversare la membrana mitocondriale interna 8 Sistemi navetta di substrati: si servono di isoforme citoplasmatiche e mitocondriali dello stesso enzima per trasportare gli elettroni dal citosol al mitocondrio. 9 Gli elettroni del NADH vengono trasportati dal citosol all’interno del mitocondrio mediante un processo che implica 3 tappe. Il sistema navetta del glicerofosfato trasferimento di una coppia di e- glicerolo 3-P deidrogenasi isozima FAD dip. 10 Shuttle malato – aspartato (fegato, muscolo cardiaco) il NADH(H+) formatosi nella glicolisi può essere ossidato a spese dell’ossalacetato: (e-) 4 1 malato deidrogenasi (reaz. di riduzione) mmi 3 2 reaz. di transamminazione malato deidrogenasi ossidato nella catena respiratoria (reaz. di ossidazione) 11 MITOCONDRIO Meccanismo conformazionale del traslocatore ADP-ATP: adenina nucleotide traslocasi 12 I due sistemi di trasporto mitocondriale per il Ca 2+ I MT funzionano da tamponi per il calcio citosolico. Importanza della distribuzione intracellulare del calcio: contrazione muscolare, trasmissione neuronale, secrezione, azione degli ormoni antiporto con ioni Na+ L’entrata nella matrice è promossa dal potenziale di membrana negativo che attrae gli ioni positivi. La velocità dipende dalla [Ca++] esterna matrice mitocondriale Se la [Ca++]cit Se la [Ca++] cit = aumenta l’entrata = diminuisce l’entrata 13 La catena respiratoria libera energia grazie alla differenza dei potenziali di riduzione tra il donatore (NADH + H+) e l’accettore (O2) di elettroni. Gran parte di questa energia viene impiegata dall’ATP sintasi per convertire l’ADP in ATP 14 Il flusso di elettroni può produrre un lavoro biologico: nel “circuito che potenzia un motore”, la fonte di e- può essere una batteria che contiene due specie chimiche che hanno una diversa affinità per gli e- . Il flusso elettronico procederà spontaneamente nel circuito guidato da una forza proporzionale alla differenza tra le due affinità detta forza elettromotrice (fem). La fem può generare un lavoro se al circuito viene collegato un trasduttore energetico. 15 In un analogo “circuito biologico” la fonte di elettroni è un composto ridotto come per es. il glucosio. Quando viene ossidato enzimaticamente sono rilasciati e- che fluiscono spontaneamente attraverso una serie di trasportatori intermedi fino a raggiungere una specie chimica con un’elevata affinità per gli e- come l’ossigeno. La forza motrice che si genera fornisce energia ad una varietà di trasduttori molecolari (enzimi e proteine) che compiono un lavoro biologico. 16 L’ossidazione biologica completa di sostanze organiche nutrienti crea un flusso di elettroni che procede verso l’ossigeno mediante una serie di trasportatori di elettroni intermedi che si comportano come un ponte a cestelli: reazione redox esoergonica: gli elettroni passano spontaneamente da un trasportatore a bassa affinità per gli elettroni ad un altro con affinità maggiore e così via. La reazione complessiva è: sostanza organica + O2 D CO2 + H2O + energia 17 Il sistema mitocondriale di trasporto degli e- non è altro che una sequenza di reazioni di ossido riduzione Durante le reazioni redox, gli elettroni direttamente dai donatori agli accettori: trasferiti possono passare Fe2+ + Cu2+ D Fe3+ + Cu+ In alternativa, gli elettroni possono essere trasferiti da un donatore a un accettore mediante un trasportatore di elettroni: XH2 + Y D X +Y H2 NADH + H+ + FAD NAD+ + FADH2 18 Le forme ossidate e ridotte dei composti sono dette coppie redox. donatore di e- (agente riducente) accettore di e- (agente ossidante) La facilità con la quale gli e- passano dal riducente all’ossidante è espressa quantitativamente dal valore del potenziale di ossido-riduzione del sistema. 19 donatore di e- (agente riducente) accettore di e- (agente ossidante) In una coppia redox il riducente con un potenziale di ox-red molto negativo cede facilmente i suoi e- a coppie con potenziali redox meno negativi o più positivi Viceversa un forte ossidante (caratterizzato da un potenziale redox molto positivo) mostra una grande affinità per gli e-. 20 Conoscendo i potenziali standard di ox-red si può predire la direzione del flusso di e- Il più forte agente ossidante cioè ha elevata affinità per gli elettroni e- 21 La catena di trasporto degli elettroni può essere divisa in 4 complessi: Schema dei complessi e del percorso degli elettroni nella catena di trasporto degli elettroni nei mitocondri 22 La catena respiratoria comprende 3 complessi proteici inseriti nella membrana mitocondriale interna (complessi I, III e IV) e molecole di trasferimento mobili, quali il coenzima Q e il citocromo-c: • complesso I: NADH deidrogenasi • complesso III: citocromo-c reduttasi • complesso IV: citocromo-c ossidasi II Il complesso II è la succinato deidrogenasi, è l’unico enzima presente del ciclo dell’acido citrico presente nella membrana interna mitocondriale. Il complesso V, l’ATP sintasi, non 23 partecipa al trasferimento di elettroni Tutti i complessi della catena respiratoria sono formati da numerose subunità polipeptidiche e contengono una serie di coenzimi redox legati alle proteine: • Flavine: FMN e FAD nei complessi I e II • Coenzima Q • Centri Ferro-Zolfo: complessi I, II e III • Gruppi eme: II, III e IV I trasportatori flavinici e il CoQ sono trasportatori misti: trasportano elettroni e H+ I centri Ferro-Zolfo e i gruppi eme dei citocromi sono trasportatori puri: trasportano elettroni 24 Nicotinamide adenina dinucleotide (NAD+) e nicotinamide adenina dinucleotide fosfato (NADP+). R= H nel NAD+ R= PO3–- nel NADP 25 • L’FMN e il FAD subiscono delle reazioni di ossido-riduzione reversibili. L’FMN è saldamente legato al complesso-I della catena respiratoria e non si comporta come un substrato diffusibile. Il FAD è tenacemente legato al complesso II, o succinato deidrogenasi. Le flavine sono dei derivati della vitavina riboflavina 26 Flavina adenina dinucleotide (FAD) Costituita da FMN + AMP 27 •Il CoQ è diffusibile e può spostarsi, durante il trasporto elettronico, tra molecole donatrici e accettrici. 28 Stati di ossidazione di Flavin mononuleotide (FMN) e Coenzima Q (CoQ) o ubichinone IL FMN e il CoQ (che possono trasferire uno o due elettroni per volta) sono il punto di contatto tra il donatore di due elettroni NADH e i citocromi accettori di un solo elettrone 29 Forma ossidata Forma radicalica Forma ridotta 30 • I centri Fe-S, come i nucleotidi flavinici, sono strettamente legati alle proteine. •Il ferro è legato allo zolfo elementare e ai gruppi tiolici di cisteine messe a disposizione dalla proteina. •Benchè in un centro possano essere presenti vari atomi di ferro, i centri ferro-zolfo partecipano a reazioni di trasferimento di un solo elettrone Esempi di centri ferro-zolfo 31 • I citocromi rappresentano l’ultima classe di componenti che partecipano al trasporto degli elettroni. •I citocromi sono delle proteine contenenti eme. •Ciascun citocromo è costituito da una catena polipeptidica e da un gruppo eme specifico. •Il ferro presente nei citocromi, ma non quello nell’emoglobina, subisce delle ossidoriduzioni fisiologiche passando dallo stato ferroso (2+) a quello ferrico (3+) e viceversa Struttura del citocromo c mitocondriale 32 Esempi di gruppi eme I gruppi eme sono composti tetrapirrolici contenenti ferro 33 Sequenza del trasporto degli elettroni 4 H+ Complesso I Complesso II Succinato deidrogenasi-FAD FeS, cit b560 34 Complesso I (NADH deidrogenasi) NADH Q NADH deidrogenasi (FMN) 4 H+ 4 H+ 35 Complesso II: catalizza l’ossidazione del FADH2 da parte del CoQ FADH2 + CoQ (ossidato) FAD + CoQ (ridotto) 4 H+ Complesso I Complesso II Succinato deidrogenasi/ a glicerofosfato deidrogenasi 36 Il complesso III catalizza l’ossidazione del CoQ (ridotto) da parte del citocromo c CoQ (ridotto) + citocromo c citocromo c (ridotto) + CoQ (ossidato) (ossidato) 4 H+ Complesso I Complesso II 37 Ciclo Q : 1) accoppiamento del trasferimento degli e(dal CoQ al cit.c) con il trasporto dei protoni transmembrana. 2) facilita il passaggiio dall’ubichinolo (che trasporta 2 e-) al cit.c (che trasporta 1 e-) (*) (*) il semichinone trasferisce l’e- prima ad un centro FeS detto Centro di Rieskie e poi al cit.c1. 38 Ciclo Q: 1) e- centro FeS ridotto cit. c1 cit. c (*) 2) e- cit. bL cit. bH UQ (ubichinone ossidato) ubichinone ridotto (semichinonico) che capta due protoni dal versante della matrice per formare QH2 39 40 Il complesso IV (citocromo c-ossidasi) catalizza l’ossidazione del citocromo c ridotto da parte dell’O2, l’accettore terminale degli elettroni nel processo di trasporto degli elettroni citocromo c (ridotto) 4 H+ 2 H+ Complesso I citocromo c (ossidato) + H2O + ½ O2 Complesso II citocromo c ossidasi 41 42 43 Anemia da carenza di Ferro Fabbisogno di ferro in un uomo adulto: 1mg/die in una donna in età fertile: 2mg/die in una donna in gravidanza: 3mg/die Il ferro è necessario per il mantenimento di una quantità normale di Hb, di citocromi e di centri ferro-zolfo. 44 45 46 47 48 Microfotografia elettronica di particelle submitocondriali sulla membrana mitocondriale interna. 49 Struttura del complesso mitocondriale dell’ATP sintasi 50 51 L’ENERGIA LIBERA A B DG= GB - GA il cambiamento di energia libera è uguale alla differenza delle energie libere di A e B Se la [B] è più elevata della [A], la reazione A è favorita, è spontanea, è esoergonica: DG0 Se la [A] è più elevata della [B], la reazione A è sfavorita, non spontanea, endoergonica: DG0 B 0 (energia libera negativa) B 0 (energia libera positiva) 52 Il trasferimento di energia dai coenzimi ridotti ai legami fosforici ad alta energia NADH + H+ + ½ O2 NAD+ + H2O DG°’= -220 kJ/mol (-52 kcal/mol) ADP + Pi ATP + H2O DG°’= -30,5 kJ/mol (-7,3 kcal/mol) 53 Il dinitrofenolo, farmaco dimagrante 54 Proteine disaccoppianti UCP, UnCoupling Proteins UCP 1 (termogenina): presente nel tessuto adiposo bruno UCP 2:ubiquitaria UCP 3: muscolo scheletrico UCP 4,UCP 5: cervello 55 56 57 58 Regolazione della fosforilazione ossidativa regolazione retroattiva: ADP stimola la respirazione e la sintesi di ATP ATP le inibisce regolazione mediata da modifiche covalenti: il complesso IV fosforilato è inattivo (PKA) e defosforilato è attivo (fosfatasi) 59 Nella riduzione di O2 si cela un pericolo: -il trasferimento di 4 elettroni dà origine a prodotti innocui - il trasferimento di un singolo elettrone forma un anione superossido -il trasferimento di 2 elettroni genera un perossido O2 e- . O2 - e- O2 2- 60 I derivati tossici dell’ossigeno molecolare (ROS) vengono rimossi da enzimi protettivi superossido dismutasi . 2 O2 + 2H+ . 2 H2O Catalasi 2 GSH + H2O2 O2 + H2O2 O2 + 2H2O GSSG + 2 H2O 61 Miopatie mitocondriali -il lattato si accumula nel liquido cerebrospinale -crampi, debolezza muscolare, encefalopatia -esame dei MT del tessuto muscolare -trattamento con CoQ10 -Carenza di CoQ10: forma rara che colpisce i bambini. -Neuropatia ottica ereditaria di Leber (mutazione del Complesso I) -I mitocondri svolgono un ruolo centrale nell’apoptosi 62
Scaricare