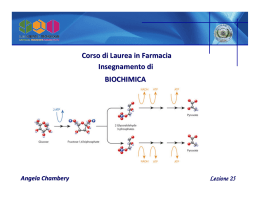
GLICOLISI e CATABOLISMO degli ESOSI pH= 7.0 Digestione dei polisaccaridi Amilasi salivare e pancreatica = endoglucosidasi Digestione degli oligosaccaridi maltosio, maltotriosio, destrine, Saccarosio, lattosio. Saccarasi, lattasi, maltasi, isomaltasi dell’intestino Deficienza congenita o acquisita delle disaccaridasi intestinali: 1. DEFICIENZA ASPECIFICA 2. DEFICIENZA DI LATTASI nell’adulto nel lattante (intolleranza al lattosio) Carrier per il glucosio. Le cellule dell’epitelio intestinale assumono il glucosio proveniente dalla dieta mediante un simporto dipendente dal sodio. Modello di un trasportatore del glucosio nei mammiferi Ciascun membro di questa famiglia è costituito da una singola catena polipeptidica lunga circa 500 residui Km= 1mM (alta affinità per il glucosio) [glucosio]siero = 4-8mM (insulino-indipendente) Km= 1mM (alta affinità per il glucosio) [glucosio]siero = 4-8mM Nel fegato il glucosio viene assunto con un sistema insulino-indipendente realizzato dal trasportatore Glut-2. , pancreas Km= 15-20mM Il glucosio entra solo quando la [glucosio]ematico è alta. Quando il glucosio entra nelle cellule b del pancreas lo induce a sintetizzare insulina. Km= 5mM GLUT4 nel muscolo e nelle cellule adipose L’assunzione di glucosio da parte del muscolo e delle cellule adipose viene regolata dall’esocitosi insulino-dipendente di vescicole membranose contenenti GLUT4. Con la rimozione dell’insulina, il processo si inverte in un processo di endocitosi La glicolisi è la sequenza di reazioni che metabolizza una molecola di glucosio a due di piruvato con la produzione netta di due molecole di ATP. Questo processo è anaerobico condizioni aerobiche condizioni anaerobiche nel lievito Reazione irreversibile La molecola del glucosio induce un cambiamento conformazionale nell’esochinasi. Esochinasi Esochinasi: inibita dal glucosio 6-P Isoenzima tipo I: nel cervello (attivato dal Pi) Isoenzima tipo II: nel muscolo Deficienza ereditaria di esochinasi (anemia emolitica) Nel fegato sono presenti entrambi gli enzimi. Glucochinasi 1. Presente nel fegato 2. Specificità per il glucosio 3. Bassa affinità 4. Inducibile (dieta, insulina) Il metabolismo glucidico è importante per gli eritrociti • nel deficit di esochinasi le concentrazioni di tutti gli intermedi glicolitici sono basse e di conseguenza la concentrazione di 2,3bifosfoglicerato è anch’essa diminuita mentre l’affinità dell’emoglobina per l’ossigeno risulta aumentata (curva verde) • nel deficit di piruvato chinasi si ha un aumento nella concentrazione di 2,3bifosfoglicerato con diminuzione dell’affinità dell’emoglobina per l’ossigeno (curva blu) SEGUE Isomerizzazione di un esosio in un chetosio Fosfoglucosio isomerasi II tappa di utilizzo dell’ATP (reazione irreversibile) Fosfofruttochinasi Svolge un ruolo centrale nel controllo della glicolisi Demolizione dell’esoso in triosi Aldolasi Gliceraldeide 3-P e diidrossiacetone fosfato sono interconvertibili Trioso fosfato isomerasi (enzima perfetto, mantiene le concentrazioni in equilibrio). Reazione accoppiata: l’energia derivata dall’ossidazione dell’aldeide è utilizzata per la formazione del legame fosfo-anidridico. (Primo intermedio ad alta energia) Gliceraldeide 3-P deidrogenasi 1 molecola di glucosio è stata trasformata in 2 molecole di GAP. Sono stati consumati 2 ATP (investimento di energia). Ia fosforilazione dell’ADP Fosfoglicerato chinasi Trasferimento intramolecolare del radicale fosforico Fosfoglicerato mutasi Formazione del PEP Enolasi Reazione irreversibile II fosforilazione dell’ADP Piruvato chinasi Glucosio + 2 NAD+ + 2 ATP + 2 Pi 2 piruvato + 2 NADH + 2 ATP + 2 H2O + 4 H+ 1. ATP. Per ogni molecola di glucosio: investimento di 2 ATP (fase I) sintesi di 4 ATP (fase II) Resa netta 2 ATP 2. NADH. Fonte di energia libera che può essere recuperata mediante successiva ossidazione. In condizioni di aerobiosi: NADH (e-) trasportatori di e- Sintesi di ATP (fosforilazione ossidativa Rigenerazione di NAD+ 3. PIRUVATO O2 Fermentazione: il destino anaerobico del del piruvato In condizioni aerobiche: piruvato CO2 + H2O In condizioni anaerobiche: piruvato Via della fermentazione Fermentazione omolattica (muscolo) COO- NADH + H+ COO- NAD+ O HO CH3 Acido piruvico lattico deidrogenasi H CH3 Acido lattico Il lattato ottenuto può essere riconvertito in glucosio dal fegato nella fase di recupero dopo lo sforzo Fermentazione alcolica I lieviti (saccharomyces cerevisiae) e altri microorganismi fermentano il glucosio ad etanolo e CO2 invece che a lattato, grazie alla presenza della piruvato decarbossilasi e dell’alcol deidrogenasi COO- O CO2 NADH + H+ O CH3 H3C piruvato decarbossilasi, TPP H NAD+ CH3CH2OH alcol deidrogenasi cellule di saccharomyces cerevisiae in gemmazione Le due reazioni della fermentazione alcolica Tiamina pirofosfato Meccanismo di reazione della piruvato decarbossilasi Deficienza di tiamina (vit. B1): beriberi 1. Attacco nucleofilo della TPP al carbonio carbonilico del piruvato. 2. Rilascio di CO2 (TPP=trappola per gli elettroni) 3. Protonazione del carbanione 4. Eliminazione della TPP e rilascio del prodotto. Bilancio energetico della glicolisi: TAPPA ATP 1) Glucosio G-6-P -1 2) F-6-P F-1,6-dP -1 3) 1,3 BPG 3-PG +2 4) PEP Piruvato +2 ______ netto + 2 Efficienza della glicolisi aerobica TAPPA ATP 1) Glucosio 2 Piruvato +2 2) 2 Piruvato 6 CO2 + 30 3) 2 NADH (H+) 2 NAD+ Glicolisi citoplasmatica Processi mitocondriali +6 ______ netto + 38 La velocità del processo è regolata da 2 enzimi: fosfofrutto chinasi (PFK-1) piruvato chinasi Entrambi gli enzimi sono regolati allostericamente La 6-fosfofrutto-1cinasi è il principale sito di regolazione della glicolisi E’ un enzima tetramerico caratterizzato da due stati conformazionali, R e T. L’ATP si comporta sia da substrato sia da inibitore allosterico Ogni subunità della PFK ha due siti di legame per l’ATP: Uno è il sito per il substrato (con eguale efficienza in entrambe le conformazioni) Il sito dell’inibitore lega l’ATP esclusivamente nello stato T . L’ altro substrato della PFK, il F6P, si lega allo stato R. Un’alta concentrazione di ATP agisce da inibitore allosterico,spostando l’equilibrio T R verso T con diminuizione dell’affinità della PFK per il F6P. PFK Attivatori: AMP, F2,6P Inibitore: ATP, pH acido, citrato La glicolisi è stimolata quando la carica energetica diminuisce (ATP/AMP) ADP + ADP ATP + AMP AMP: segnale per lo stato a bassa energia adenilato chinasi Regolazione particolarmente sensibile da parte dell’AMP [ATP] + [ADP] + [AMP] = K [ATP] [ADP] [ADP] [AMP] Amplificazione: Piccole variazioni nella [ATP] generano grandi variazioni nella [AMP] Regolazione allosterica della fosfofruttochinasi da parte dell’ATP pH La sensibilità della 6PFK agli ioni H+ è un meccanismo di difesa contro l’acidosi lattica (diminuizione del pH ematico) insieme al simporto acido lattico-H+ Citrato Un’elevata concentrazione di citrato indica che i precursori biosintetici sono abbondanti e non deve essere degradato altro glucosio (indispensabile per il cervello). PFK2 FBP2 Struttura dei domini dell’enzima bifunzionale fosfofruttochinasi 2 PFK2 e FBPasi sono presenti in una singola catena polipeptidica 2 3 1 Sequenza amminoacidica dell’enzima Regolazione della sintesi e della degradazione del fruttosio 2,6-bisfosfato (isoforma L, epatica) [glucosio] Una diminuzione del glucosio ematico segnalato dal glucagone, determina la fosforilazione dell’enzima bifunzionale con coseguente diminuzione del fruttosio 2,6-bifosfato. Unaumento della concentrazione di fruttosio 6fosfato accelera la formazione di fruttosio 2,6.bifosfato facilitando la defosfrilazione dell’enzima bifunzionale 2 o 1 Fegato e cuore contengono differenti isoenzimi dell’enzima bifunzionale PFK/FBPasi che danno differente risposta allo stesso ormone (adrenalina). adrenalina glucagone Fegato: (inibizione della glicolisi) Cuore: (attivazione glicolisi) Anche la piruvato cinasi è un enzima glicolitico soggetto a regolazione Piruvato chinasi - ATP - Alanina e precursori biosintetici + F1,6BP 2 isoenzimi: L (nel fegato) ed M (nel muscolo). Piruvato cinasi (tipo L) è soggetto a regolazione mediante fosforilazione (per impedire al fegato di consumare glucosio quando l’encefalo e i muscoli hanno maggiore necessità di questo composto). 1 Le principali caratteristiche della regolazione della glicolisi Alterazione dell’espressione genica nei tumori causata dall’ipossia Le cellule cancerose crescono più rapidamente dei vasi sanguigni che le nutrono • in assenza di ossigeno l’ HIF-1 ( fattore di trascrizione inducibile dall’ipossia ) aumenta l’espressione della maggior parte decli enzimi glicolitici e dei trasportatori del glucosio GLUT-1 e GLUT-3 • la captazione del glucosio è collegata alla aggressività del tumore • l’HIF-1 stimola la crescita di nuovi tumori aumentando l’espressione delle molecole segnale, come il fattore di crescita vascolare endoteliale ( VEGF ), che facilita la crescita dei vasi sanguigni Oltre al glucosio molti altri carboidrati possono entrare nella via glicolitica e subire degradazioni che liberano energia; i più importanti sono: • polisaccaridi (amido, glicogeno) • disaccaridi (maltosio, lattosio, saccarosio) • altri monosaccaridi (fruttosio, mannosio, galattosio) Entrata nella glicolisi di altri esosi Punti d’ingresso nella glicolisi per galattosio e fruttosio GLUCOSIO (via gluconeogenesi) Glicerolo fosfato deidrogenasi ACIDO LATTICO (via fruttolisi) Grassi Diidrossiacetonfosfato L’uomo consuma considerevoli quantità di fruttosio sotto forma di saccarosio ( glucosio+fruttosio) Gran parte del fruttosio ottenuto nell’intestino tenue dall’idrolisi del saccarosio è convertito in glucosio La carenza di fruttochinasi è responsabile dell FRUTTOSURIA ESSENZIALE una anomalia metabolica benigna La carenza del fruttosio 1-fosfato-aldolasi è responsabile della INTOLLERANZA EREDITARIA AL FRUTTOSIO Metabolismo del galattosio galattosemia GALATTOSEMIA • Gli individui che hanno questo difetto sono incapaci di metabolizzare il galattosio proveniente dal lattosio (zucchero del latte ) a metaboliti del glucosio, spesso con conseguente formazione di: cataratta arresto dell’accrescimento ritardo mentale eventuale morte da danno epetico • Il disturbo genetico è causato da una carenza cellulare di galattocinasi con disordine lieve, o di galattosio 1-fosfato uridiltrasferasi che causa la malattia più grave. FINE
Scaricare