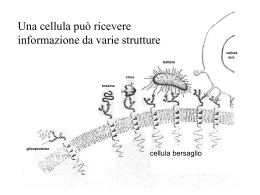
Metabolismo • Rappresenta il consumo energetico di un individuo • Consumo energetico = consumo di base (a riposo) + consumo necessario per le attività quotidiane • Il fabbisogno energetico viene soddisfatto dall’energia che viene liberata dalla metabolizzazione dei legami chimici dei glicidi, lipidi, protidi assunti giornalmente • In caso di necessità, l’organismo utilizza le sostanze che sono immagazzinate sotto forma di riserva, mobilizzandole e metabolizzandole Condizioni di sazietà e di digiuno • A seconda del tempo trascorso dall’ultima introduzione degli alimenti, possiamo distinguere due condizioni: – Stato di sazietà o di assorbimento = periodo successivo al pasto: gli alimenti vengono assorbiti, usati, accumulati – Stato di digiuno o postassorbimento = i nutrienti non sono più presenti in circolo: l’organismo utilizza le riserve accumulate Stato di digiuno o fase catabolica • Tutti i depositi (grassi, zuccheri, proteine) vengono ridotti allo scopo di mantenere la concentrazione ematica di glucosio entro valori sufficienti ad apportare energia al SNC Fonte principale ma poco duratura Il muscolo manca dell’enzima necessario a trasformare il glucosio-6- P in glucosio, quindi forma piruvato (condizioni aerobie) o lattato (c. anaerobie) che invia al fegato Adattamenti dell’organismo al digiuno prolungato • Riduzione del 15-20% della spesa energetica totale • Riduzione del consumo di glucosio • Aumento dell’utilizzo dei corpi chetonici nel SNC • Riduzione della gluconeogenesi • Riduzione della degradazione proteica a 25-30 gr/die • Riduzione del peso di c.a 300 gr/die (2/3 a carico dei grassi e 1/3 a carico delle proteine) Destino dei nutrienti in condizioni di sazietà (fase anabolica) • Il destino principale dei carboidrati che vengono assorbiti principalmente come glucosio: – Subito metabolizzati e l’energia liberata viene intrappolata tramite le vie aerobie in legami ad alta energia (ATP,…) • Delle proteine assorbite principalmente come aminoacidi: – Utilizzati per la sintesi delle proteine, componenti necessari alla crescita e mantenimento di cellule e tessuti • Dei lipidi, assorbiti principalmente come trigliceridi: – Accumulati come glicogeno e lipidi Interdipendenza del metabolismo dei diversi nutrienti Assorbiti come Assorbiti come Assorbiti come Importanti nel digiuno Energia Energia Energia Le vie biochimiche per la produzione di energia si intersecano in vari punti, in ciascuno dei quali gli enzimi vengono controllati • La regolazione di tutte le vie metaboliche che portano agli adattamenti necessari alle condizioni di digiuno e di sazietà è sotto il controllo endocrino, integrato da un controllo di tipo nervoso Il sistema endocrino • Funzione del sistema endocrino è quella di mantenere la stabilità dell’ambiente interno a fronte di variazioni provenienti dall’ambiente esterno ( cambiamenti di temperatura, assunzione di elementi nutritivi, ecc.) Relazioni tra sistema nervoso e sistema endocrino Stimoli simili sono capaci di evocare l’attività di entrambi i sistemi. Gli ormoni secreti dalle cellule endocrine e trasportati per via ematica sono analoghi ai neurotrasmettitori liberati dai neuroni dopo essere stati trasportati lungo gli assoni. I neurotrasmettitori possono anche stimolare la liberazione di ormoni o agire essi stessi come ormoni. In entrambi i sistemi le risposte vengono mediate da recettori e possono consistere in variazioni sia fisiche sia chimiche. Modalità di distribuzione degli ormoni Meccanismi di segnali endocrini e nervosi Le molecole ormonali vengono trasportate dal sangue in forma libera (ormoni idrosolubili) o legate a proteine plasmatiche (ormoni liposolubili. Le molecole biologicamente attive sono quelle libere e che si liberano dalle proteine trasportatrici. Agiscono solo sulle cellule che hanno i recettori specifici per quell’ormone (cellule bersaglio). Nelle cellule nervose i neurotrasmettitori sono rilasciati dalle terminazioni assoniche a seguito dell’arrivo di impulsi elettrici e si legano ai recettori postsinaptici. Elementi costitutivi del sistema endocrino • Ghiandole o sistemi cellulari diffusi • Ormoni (sostanze che danno effetti a concentrazioni molto basse e, in genere, a distanza dal luogo di produzione) • Tessuti bersaglio (con siti recettoriali specifici per il proprio ormone ospite = complesso ormone-recettore. I due elementi devono presentare un alto grado di complementarietà e possono presentarsi secondo due schemi che rispondono al principio della “chiave e della sua serratura” o a quello della “chiusura lampo” a più siti) • Prodotti del tessuto bersaglio (che modificano l’attività sia di altri organi, sia delle stesse strutture endocrine da cui è partito il messaggio ormonale) Ghiandole e sistemi cellulari • Le cellule possono essere raggruppate ed organizzate in ghiandole (ipofisi, tiroide, paratiroidi, isole di Langherans, corticale e midollare del surrene, epifisi, timo, testicoli, ovaie) • oppure possono essere “diffuse” ed inserite all’interno delle pareti degli organi o nei tessuti (nei nuclei sopraottico e paraventricolare dell’ipotalamo, cellule sparse ipotalamiche, cellule endocrine del sistema gastroenterico, ecc) • Alcune ghiandole sono sotto il controllo del sistema ipotalamo-ipofisario (tiroide, surrene, ghiandole sessuali), altre ne sono indipendenti (paratiroidi, pancreas endocrino) Sedi delle principali ghiandole endocrine Gli ormoni • Modificano la velocità dei processi chimici intracellulari • Agiscono in concentrazioni minime • Hanno breve vita in circolo Ormoni: classificazione • Ormoni proteici: (GH, ADH, ecc) costituiti da amminoacidi, sintetizzati nel reticolo endoplasmatico rugoso, derivano da una molecola più lunga (preprormone) • Ormoni glicoproteici: (TSH, LH, ecc) simili ai precedenti, vengono sintetizzati per la parte proteica nel reticolo endoplasmatico e poi assemblati con la parte glicidica nell’apparato di Golgi • Ormoni steroidei: (o. sessuali, aldosterone, ecc) derivati dal colesterolo • Ormoni derivati dalle amine: distinti in ormoni derivati dalla tirosina direttamente (ormoni tiroidei) o attraverso la diidrossifenilalanina o Dopa (catecolamine) e derivati dal triptofano (melatonina) Esempi dei vari tipi di ormoni Vie di sintesi degli ormoni steroidei Caratteristiche dei diversi tipi di ormoni • Gli ormoni peptidici (proteici e glicoproteici) sono considerati ormoni di tipo I • Gli ormoni steroidei sono considerati ormoni di tipo II • Gli ormoni amminici sono considerati di tipo intermedio in quanto presentano in parte le caratteristiche del I tipo (sintesi) e in parte quelle del II tipo (liberazione e trasporto) Caratteristiche degli ormoni peptidici, es. insulina • Vengono sintetizzati in eccesso ed accumulati in vescicole secretorie • Vengono rilasciati per esocitosi • Sono idrosolubili e quindi circolano liberamente disciolti nel plasma • Hanno emivita breve (da pochi minuti a una o poche ore) • Hanno il recettore sulla membrana cellulare • Agiscono attivando un sistema di secondi messaggeri nella cellula-bersaglio • La cellula bersaglio risponde modificando proteine già esistenti • La degradazione avviene tramite peptidasi renali, plasmatiche o tramite enzimi delle cellule bersaglio Caratteristiche degli ormoni steroidei, es. cortisolo • Vengono sintetizzati da precursori su domanda • Vengono rilasciati per diffusione semplice • Sono liposolubili e quindi circolano nel plasma legati a proteine di trasporto (albumina, globuline) • Hanno emivita lunga (da poche ore a qualche giorno) • Hanno il recettore nel citoplasma o nel nucleo. Alcuni lo hanno sulla membrana cellulare • Agiscono attivando nella cellula-bersaglio i geni per la trascrizione e traduzione • La cellula bersaglio risponde inducendo la sintesi di nuove proteine • La degradazione avviene tramite enzimi epatici per riduzione e coniugazione Le prostaglandine • Derivano dall’acido arachidonico (liberato dai fosfolipidi di membrana ad opera della fosfolipasi A2) per azione della ciclossigenasi che lo trasforma in precursore delle prostaglandine • Sono molecole di segnale ad azione ormon-simile, prodotte da diversi tipi di cellule in vari tessuti dell’organismo • Sono liberate rapidamente dopo la sintesi, raggiungono le cellule bersaglio attraverso il sangue ed interagiscono con esse tramite un recettore membranario • Le caratteristiche strutturali e il rapido catabolismo limitano i loro effetti in un ristretto raggio d’azione, per cui alcuni AA li considerano ormoni paracrini • Hanno diverse funzioni, tra cui: partecipano alla regolazione della motilità gastrointestinale, e al trasporto ionico a livello dei tubuli renali, regolano la liberazione di mediatori del SN, possono stimolare la contrazione o il rilasciamento del muscolo liscio, possono facilitare vasodilatazione o vasocostrizione nelle arteriole, sono importanti mediatori dell’infiammazione….. Attività ormonale • L’attività delle cellule endocrine è sotto l’influenza di stimoli eccitatori (in rosso) ed inibitori (in blu) di natura ormonale, umorale, nervosa • L’ormone liberato determina una risposta nella cellula bersaglio • Tale risposta regola l’attività di biosintesi e secrezione della cellula endocrina con un meccanismo retroattivo, più frequentemente negativo (feedback negativo) • Anche la concentrazione plasmatica dell’ormone libero può agire con un meccanismo a feedback sulla regolazione della stessa cellula endocrina che lo ha prodotto Alterazioni dell’attività ormonale • A carico della cellula endocrina: es. maggiore o minore secrezione • A carico della cellula target: es. variazione di densità dei recettori, variazione di affinità e/o capacità di riconoscere l’ormone I recettori della cellula target Qualsiasi composto che si leghi ad alta specificità al recettore viene denominato ligando. Se il ligando evoca una risposta fisiologica viene denominato agonista. Se il ligando si lega con alta specificità al recettore ma non evoca alcuna risposta, viene detto antagonista perché occupa i recettori interferendo con l’azione degli agonisti La concentrazione di molti recettori può essere regolata • Up-regulation: aumento della risposta recettoriale per ipersensibilizzazione • Down-regulation: diminuzione della risposta recettoriale per desensibilizzazione dovuta a prolungata esposizione delle cellule target ad alte concentrazioni ematiche ormonali. • Per quanto riguarda il numero di recettori sulla membrana plasmatica spesso diminuisce (Down-regulation ) perché il legame con molecole agoniste stimola l’internalizzazione dei complessi recettori-agonisti per un processo di endocitosi, seguito al loro trasferimento nei lisosomi e conseguente degradazione. • Per quanto concerne i recettori intracellulari il meccanismo è meno chiaro Gli ormoni steroidei: la sintesi • Vengono sintetizzati a partire dal colesterolo, che si trova nelle ghiandole endocrine in vacuoli di deposito sotto forma di esteri del colesterolo • Gli esteri del colesterolo derivano o dalla sintesi endogena, intracellulare, di colesterolo a partire dall’AcetilCoA, o dal colesterolo esogeno che viene trasportato alle cellule dalle lipoproteine ( in particolare le LDL) attraverso il circolo ematico • La sintesi avviene nel reticolo endoplasmico liscio e nei mitocondri • Gli ormoni steroidei non vengono accumulati, per cui tutti gli ormoni sintetizzati vengono secreti. Gli ormoni steroidei: secrezione • Gli ormoni vengono liberati in dosi sempre molto basse (all’incirca 10-10 mol/l) • La liberazione degli ormoni può avvenire in maniera acuta (secondo necessità), o con una dinamica su base ritmica (24h, di meno, di più, mensile…) • La liberazione avviene per semplice diffusione attraverso la membrana plasmatica Gli ormoni steroidei: il trasporto • Il trasporto avviene tramite alcune proteine di trasporto circolanti specifiche o aspecifiche (albumine) • Le proteine servono anche a proteggere l’ormone dall’inattivazione • Quindi l’emivita degli ormoni steroidei è più lunga di quella degli ormoni proteici • La presenza delle proteine di trasporto consente una sorta di “deposito” in circolo • Per essere utilizzato l’ormone deve staccarsi dalla proteina trasportatrice, cioè deve trovarsi in forma libera • La forma libera è l’unica biologicamente attiva • Tra la forma libera e quella legata c’è una sorta di equilibrio tale che, man mano che la forma libera viene utilizzata, una quota equivalente viene ceduta dalla proteina trasportatrice Proteine di trasporto circolanti Meccanismo di azione cellulare degli ormoni steroidei Risposte lente Recettori degli ormoni steroidei e tiroidei Gli ormoni steroidei si legano a recettori citoplasmatici, mentre quelli tiroidei raggiungono direttamente il nucleo e si legano a recettori nucleari L’ormone steroideo raggiunge il recettore citoplasmatico legato a molecole di accompagnamento (tra le quali le proteine dello shock termico – HSP) L’ormone si lega al recettore alla sua estremità C-terminale e ne determina una modifica conformazionale: il complesso ormone-recettore si trasferisce nel nucleo Il complesso ormone-recettore dimerizza Il dimero recettore-ormone va ad attivare sequenze di DNA chiamate elementi di HSP risposta ormonale (HRE) Di norma si ha aumento di trascrizione genica, ma in alcuni casi si può avere una regolazione negativa con repressione della trascrizione Si può avere repressione di trascrizione anche in caso di recettori nucleari che non legano gli ormoni HRE Effetti sull’espressione genica e sui possibili modi in cui la funzionalità cellulare può venire modificata Esempio d’azione di ormone steroideo: l’aldosterone Aumenta il trasporto di Na attraverso le cellule cambiando il ritmo di sintesi e quindi la quantità di alcune proteine chiave nella cellula Aumenta il numero dei canali al Na aperti sulla membrana cellulare (non illustrato) Sintesi di proteine della pompa Gli ormoni steroidei: eliminazione • Gli ormoni in forma libera svolgono anche un’azione di controllo a feed-back sulla ghiandola endocrina che li ha prodotti • Gli ormoni steroidei vengono attaccati da enzimi epatici con processi di riduzione e coniugazione • Gli ormoni vengono eliminati in parte per via urinaria, ma soprattutto per via fecale, grazie alla bile, dopo essere stati metabolizzati Gli ormoni peptidici Gli ormoni peptidici: la sintesi Gli ormoni proteici derivano da una molecola proteica di maggiori dimensioni detta preproormone. Da questa, per azione enzimatica, vengono staccati alcuni amminoacidi e si forma il proormone. Infine, da questo, sempre per azione enzimatica e distacco di un peptide, si forma l’ormone Tappe della sintesi degli ormoni peptidici localizzata all’estremità N-terminale I proormoni Struttura di alcuni proormoni: La parte blu rappresenta la parte attiva degli ormoni La parte rossa è quella che può far variare le caratteristiche del proormone o dell’ormone finale: es. la somatostatina può ritrovarsi nella forma 1-14 o 1-28 Gli ormoni peptidici: secrezione Uno stimolo aumenta i livelli citosolici di Ca++ e, spesso, di AMPc I granuli secretori vengono allineati e trasportati verso la membrana plasmatica grazie ad un sistema di microtubuli e microfilamenti La membrana dei granuli secretori si fonde con quella della cellula La membrana comune viene lisata, determinando la liberazione dell’ormone nello spazio interstiziale, per esocitosi Gli ormoni tiroidei non vengono depositati in granuli ben differenziati e lasciano la cellula seguendo il gradiente di concentrazione tra interno ed esterno della membrana cellulare Gli ormoni peptidici: trasporto • Circolano nel plasma non legati a proteine specifiche • Hanno emivita breve • Danno luogo ad effetti rapidi Il legame ormone-recettore • Il recettore degli ormoni proteici è localizzato sulla membrana perché la molecola non riesce ad attraversare lo strato fosfolipidico che costituisce la membrana • E’ caratterizzato da alta affinità ed alto grado di specificità, come per gli ormoni steroidei • Generalmente un recettore lega il proprio ormone specifico in maniera forte • Alcuni recettori possono legare molti altri ormoni con più bassa specificità • Il legame modifica la conformazione tridimensionale del recettore • Il legame è, di solito, prontamente reversibile I recettori • I recettori sono proteine intrinseche di membrana non fissate rigidamente ma con notevole mobilità laterale • Generalmente si tratta di glicoproteine con la porzione glicidica rivolta verso l’esterno della membrana • A seguito di aumento di concentrazione ematica dell’ormone, si può avere la saturazione dei recettori (risposta massimale dell’ormone). Successivamente, la risposta non aumenta anche se aumenta la concentrazione dell’ormone circolante • Di norma il recettore modifica la sua conformazione dopo il legame con l’ormone • Può accadere che si abbia una endocitosi dell’ormone mediata dal recettore • Il recettore funge da convertitore di segnali (trasduttore) e attiva molecole che fungono da secondi messaggeri • L’effetto è quello di dare molteplici effetti biologici che modificano la funzionalità cellulare Interazione ormone-recettore Le reazioni a cascata • Vengono innescate dal legame tra ligando e recettore • Tale legame attiva un enzima amplificatore • L’enzima amplificatore converte due o più molecole inattive A in forma attiva • Le molecole A attivate convertono due o più molecole inattive B in forma attivata e così via fino alla tappa finale in cui il substrato viene convertito nel prodotto finale • In tal modo si ottiene l’amplificazione del segnale • L’amplificazione permette di ottenere un grande effetto con un piccolo segnale e quindi con economizzazione di energia Amplificazione del segnale I recettori di membrana I recettori di membrana, attivati dal ligando, trasferiscono l’informazione all’interno della cellula (trasduzione del segnale). Le due modalità più comuni per la trasduzione del segnale sono rappresentate da: (1) apertura di canali ionici di membrana, (2) una proteinchinasi che fosforila le proteine, (3) da proteine di membrana note come proteine G. Le proteinchinasi • E’ il sistema di trasduzione più semplice, in cui una singola proteina transmembranaria funge da recettore (porzione extracellulare) e da enzima amplificatore (porzione intra cellulare) • Il legame tra ligando e recettore attiva la porzione proteica intracellulare che trasferisce un gruppo fosfato da un ATP ad una proteina plasmatica attivandola o inibendola • Poiché il fosfato viene attaccato ad un residuo di tirosina della proteina, l’enzima viene definito tirosinchinasi (questo meccanismo viene usato ad es. dall’insulina) Protein-chinasi e proteine fosforilate Le proteine fosforilate controllano uno o più dei seguenti bersagli: 1) enzimi metabolici, 2) proteine di trasporto, 3) proteine che regolano l’attività di geni e la sintesi di proteine, 4) recettori di membrana Le proteine G • Vengono così denominate perché tutte richiedono per la loro funzione GDP e GTP (guanosin di- e tri-fosfato) • L’attivazione della proteina G si verifica quando essa si lega al GTP. • La durata di questa attivazione è breve perché è la stessa attività enzimatica della proteina G a degradare il GTP a GDP • L’amplificazione ha luogo perché molte proteine G (fino a 100) sono attivate da ogni complesso ormone-recettore • Sono stati individuati più di 300 recettori accoppiati a proteine G Proteine G: meccanismi di azione La proteina G può agire in due modi : 1) apre i canali ionici di membrana, 2) stimola o inibisce un enzima amplificatore presente sul lato citoplasmatico della membrana Proteine G e apertura dei canali ionici I • L’apertura dei canali ionici in cellule attive elettricamente determina il cambiamento di concentrazione ionica nella cellula • Il movimento di ioni Na e K modifica il potenziale di membrana della cellula e crea un segnale elettrico • Il segnale elettrico agisce come II messaggero Proteine G e apertura dei canali ionici II • In cellule che non sono attive elettricamente l’ingresso di ioni può fare iniziare una risposta cellulare di altro tipo. Ad es un certo numero di recettori cellulari è connesso a canali al Ca: gli ioni Ca, una volta entrati nella cellula, agiscono come secondi messaggeri e attivano un certo numero di proteine Ca-dipendenti • Il Ca attiva le proteinchinasi come parte della cascata dei secondi messaggeri • Viene anche modificata la concentrazione del Ca citoplasmatico che regola il rilascio delle vescicole secretorie e la contrazione muscolare Apertura di canali ionici di membrana Proteine G ed enzimi amplificatori • L’attivazione o inibizione degli enzimi amplificatori costituiscono la maggior parte dei meccanismi di trasduzione del segnale • Questi enzimi inducono la sintesi dei secondi messaggeri • I secondi messaggeri attivano le proteinchinasi sia direttamente, sia tramite il rilascio di ioni Ca dai serbatoi di accumulo Enzimi amplificatori • Adenilato ciclasi: catalizza la conversione di ATP in AMPc • Guanilato ciclasi: controlla la formazione di GMPc • Fosfolipasi C: converte un fosfolipide di membrana in 2 differenti molecole che fungono entrambe da secondi messaggeri Adenilato ciclasi I cambiamenti nell’AMPc intracellulare avvengono in pochi secondi dal legame dell’ormone al recettore. Gli effetti intracellulari dell’AMPc sono molto rapidi, quindi l’ormone produce effetti acuti molto rapidi (es. glucagone, adrenalina, ACTH, LH….) Relazione tra i recettori di membrana e l’adenilato ciclasi tramite le proteine Gs e Gi • Esistono due tipi di proteine G, stimolanti ed inibenti (Gs e Gi). • Se l’ormone si lega ad un recettore di membrana Rs accoppiato ad una proteina Gs l’adenilato ciclasi viene stimolata e l’AMPc aumenta • Viceversa il legame di un ormone ad un recettore di membrana Ri accoppiato ad una proteina Gi inibisce l’adenilato ciclasi e l’AMPc diminuisce Guanilato ciclasi • Alcuni tessuti usano al posto dell’adenilato ciclasi la guanilato ciclasi • Questo enzima amplificatore controlla la formazione di GMPc a partire dal GTP • Il GMPc utilizza meccanismi d’azione analoghi a quelli dell’AMPc La fosfolipasi C • E’ un enzima amplificatore che converte il fosfatidilinositolo 4,5-bifosfato (PIP2) in inositol trifosfato (IP3) e diacilglicerolo (DG o DAG) che agiscono da secondi messaggeri • La fosfolipasi C è accoppiata funzionalmente ai recettori di membrana mediante una proteina G • Di questa proteina G è stata identificata solo una forma stimolante. Al momento non sembra che esista una forma di proteina G inibitoria per questo sistema I secondi messaggeri • I secondi messaggeri chimici conosciuti sono : • AMPc • GMPc • Diacilglicerolo (DG o DAG) • Inositolo trifosfato (IP3) • Ioni Ca++ 3’,5’-adenosinmonofosfato ciclico o AMP ciclico (AMPc) • E’ stato uno dei primi secondi messaggeri ad essere scoperto • E’ generato dall’ATP cellulare mediante l’azione dell’enzima amplificatore adenilato ciclasi, localizzato sulla superficie interna della membrana cellulare • Viene inattivato rapidamente mediante la conversione in AMP, reazione catalizzata da una o più fosfodiesterasi • Ogni aumento dell’AMPc citoplasmatico sarà di breve durata, permettendo la rapida inattivazione degli effetti da esso mediati Sistema Adenilato ciclasi-AMPc-proteine chinasi • L’AMPc agisce attivando proteine chinasi AMPcdipendenti. • Queste catalizzano la fosforilazione di poche specifiche proteine cellulari che cambiano la loro attività • Il cambiamento di attività si traduce in cambiamenti specifici nella funzione cellulare Il Guanosin monofosfato ciclico • Il GMPc utilizza meccanismi analoghi a quelli dell’AMPc, attivando proteine chinasi GMPc dipendenti che agiscono cambiando le funzioni cellulari Diacilglicerolo (DG o DAG) Inositolo trifosfato (IP3) • Il fosfatidil-inositolo è un fosfolipide componente minoritario della membrana plasmatica, localizzato principalmente sulla metà interna del doppio strato • Dopo la donazione di due gruppi P all’ATP, il fosfatidil-inositolo è convertito in fosfatidil-inositolo 4,5-bifosfato (PIP2) • Il PIP2 serve come precursore immediato per la generazione di due secondi messaggeri intracellulari, il diacilglicerolo (DG o DAG) e l’inositolo trifosfato (IP3) che sembrano agire insieme per regolare molti processi diversi • Come già detto, questa reazione è catalizzata dalla fosfolipasi C Formazione e meccanismo d’azione di IP3 e DG Meccanismo d’azione di IP3 • L’ IP3 causa il rilascio di ioni Calcio dai siti di accumulo nel reticolo endoplasmatico • Alza quindi la concentrazione del Calcio citoplasmatico libero • Il Calcio, in questo caso, agisce come terzo messaggero, andando ad influenzare diversi processi metabolici cellulari Meccanismo d’azione del DG • Il DG rimane associato alla membrana • Attiva una proteina chinasi di membrana, la chinasi C • La chinasi C catalizza la fosforilazione di proteine cellulari specifiche, alterandone l’attività • Le proteine modificate producono cambiamenti specifici nella funzione cellulare Il Calcio come secondo messaggero • Il Calcio può entrare nella cellula a seguito di diversi meccanismi: • 1) Tramite canali al calcio voltaggio-dipendenti • 2) Tramite canali controllati da un ligando • 3) Tramite canali controllati da stimoli meccanici • 4) Come già detto, può essere rilasciato dai compartimenti intracellulari ad opera di secondi messaggeri • Gli ioni Calcio nel citosol si combinano con proteine citoplasmatiche leganti il Calcio, esercitando vari effetti Effetti del Calcio sulla cellula • Il Calcio può legarsi ad alcune proteine plasmatiche ed esercitare effetti diversi: • 1) Con la Calmodulina, che si trova in tutte le cellule, modifica l’attività di enzimi o lo stato di apertura dei canali ionici • 2) Legandosi ad alcune proteine di regolazione (come la troponina) altera la funzione di proteine contrattili o di proteine del citoscheletro come i microtubuli • 3) Legandosi ad altri tipi di proteine di regolazione induce l’esocitosi di vescicole di secrezione • Il Calcio può legarsi direttamente ad un canale ionico alterandone lo stato di apertura (es. canale del K nelle cellule nervose) • L’ingresso del Calcio in un uovo fertilizzato segna l’inizio dello sviluppo dell’embrione Effetti del Calcio diretti e tramite calmodulina Sintesi degli effetti dello ione Calcio Riassunto dei meccanismi d’azione degli ormoni peptidici L’acido arachidonico • L’acido arachidonico (ac. grasso a 20 atomi di C) è prodotto dai fosfolipidi di membrana tramite l’azione dell’enzima fosfolipasi A2 (PLA2) • La PLA2 è controllata da ormoni e da altri segnali • L’ac. Arachidonico, una volta formatosi, può agire come secondo messaggero, modificando l’attività di canali ionici e di enzimi intracellulari • Ma l’ac. Arachidonico può anche essere convertito in una delle molte classi di sostanze dette eicosanoidi, molecole solubili nei lipidi, in grado di diffondere fuori della cellula comportandosi da sostanze paracrine La cascata dell’acido arachidonico I leucotrieni • Sono secreti da alcuni tipi di leucociti per azione dell’enzima lipoossigenasi sull’acido arachidonico • Hanno tre doppi legami tra gli atomi di Carbonio • Sono coinvolti nella fisiopatologia dell’asma e della reazione allergica severa nota come anafilassi • Per questo le case farmaceutiche sono da tempo impegnate per sviluppare farmaci in grado di inibire la sintesi o le azioni dei leucotrieni I prostanoidi • Sono molecole prodotte dall’azione dell’enzima cicloossigenasi (COX1 e COX2) sull’ac. Arachidonico • Includono prostaglandine e trombossani • Entrambi agiscono su diversi tessuti, inclusi il muscolo liscio, le piastrine, il rene e l’osso • In particolare, come già detto, le prostaglandine sono coinvolte nel sonno, nell’infiammazione, nel dolore e nella febbre • Farmaci antiinfiammatori non steroidei (FANS) come l’aspirina e l’ibuprofene prevengono l’infiammazione inibendo gli enzimi COX e riducendo la sintesi delle prostaglandine • Tuttavia, poiché i FANS non sono specifici possono avere effetti collaterali
Scarica