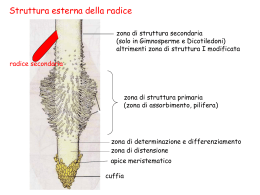
Plastidi -I cloroplasti di colore verde contenenti i pigmenti clorofilla a, b e carotenoidi (e ficobiline nei plastidi fotosintetici delle alghe rosse); -i cromoplasti di colore giallo, arancio o rosso mancano di clorofilla ma hanno carotenoidi; -i leucoplasti incolori per l’assenza di pigmenti. Come nei procarioti e nei mitocondri, i plastidi presentano una o più molecole di DNA e ribosomi deputati alla sintesi proteica. Si duplicano per scissione. Altri tipi di pigmenti si trovano nei vacuoli (viola, blu, giallo) a) Cloroplasti b) Cromoplasti c) Leucoplasti. • I cromoplasti hanno forma variabile e mancano di un sistema tilacoidale vero e proprio. Sono di colore giallo, arancio o rosso, mancano di clorofilla, ma sono ricchi in carotenoidi. Generalmente si differenziano a partire dai cloroplasti (si pensi ad esempio alla maturazione del pomodoro), ma possono derivare anche da proplastidi o leucoplasti (ad esempio nella carota). La conversione da cloroplasti comporta la degradazione delle clorofille e più in generale dell’apparato fotosintetico, e la sintesi e l’accumulo di pigmenti carotenoidi. • La conversione è legata a fattori endogeni (ormoni e nutrienti) ed ambientali (fotoperiodo e temperatura). Il processo può essere reversibile I colori vivaci conferiti dai carotenoidi a molti fiori e frutti rivestono un ruolo importante nell’attrazione di animali impollinatori e di quelli coinvolti nella dispersione dei semi. Certe volte però non è chiaro (carota, ravanello). • i flavonoidi e gli antociani. Questi metaboliti secondari sono un'ampia classe di sostanze colorate di natura fenolica che comprendono le antocianine, i flavoni, i flavonoli e gli isoflavoni. Essi danno una grande varietà di colori dal giallo fino al blu-viola e vengono accumulati nei vacuoli delle cellule • I leucoplasti sono plastidi incolori per l’assenza di pigmenti. Sono distinti in base alle sostanze prodotte e/o accumulate: - elaioplasti: accumulano lipidi; - proteinoplasti: accumulano proteine, ad esempio in alcune radici; - amiloplasti: accumulano carboidrati sotto forma di amido. Questi organelli sono particolarmente abbondanti nelle parti della pianta specializzate per l’accumulo, come radici, tuberi, rizomi, semi, midollo del fusto. I proplastidi della radice nelle cellule adulte diventeranno leucoplasti (amiloplasti):se esposta alla luce difficilmente diventeranno cloroplasti. Gli amiloplasti maturi, il cui stroma è quasi interamente occupato da granuli di amido, assumono forme e dimensioni diverse, che dipendono dalla deposizione di strati di amido intorno a un centro di formazione detto ilo, che può essere centrale o eccentrico. In alcuni casi, come nella patata, intorno all’ilo si possono osservare diversi strati concentrici più chiari e più scuri. L’amido primario che si forma durante la fotosintesi nei cloroplasti dove è deposto in piccoli granuli, durante la notte quando non si ha fotosintesi viene idrolizzato in dimeri di saccarosio (un glucosio + un fruttosio) i quali vengono poi trasferiti negli organi di riserva dove si ripolimerizzano nei leucoplasti (amiloplasti) a formare l’amido secondario costituito da amilosio ed amilopectina. L’inverdimento delle patate, causato dall’esposizione alla luce, riflette la conversione degli amiloplasti in cloroplasti. Cellule di tubero di patata Cloroplasti con amido primario • Negli organismi pluricellulari le cellule si dividono mitoticamente e si differenziano in modo da poter svolegre tutte le funzioni necessarie per la vita stessa della pianta. • Le cellule si organizzano a formare i tessuti e questi gli organi Tegumentali, Parenchimatici, Meccanici, Conduttori Foglia, Fusto, Radice organi riproduttivi Tessuti Tessuti meristematici: cell. non differenziate, mitosi Primari tessuti primari corpo primario apice vegetativo (apice fiorale), apice radicale, Secondari, es. Cambio e Fellogeno tra i tessuti definitivi dell’adulto (Gimnosperme e Dicotiledoni) tessuti secondari o definitivi Tessuti definitivi: cell. differenziate, no mitosi • Le piante hanno una CRESCITA INDEFINITA, che cioè dura per tutta la vita, anche se naturalmente la crescita è più intensa in certe stagioni dell’anno o fasi della vita del vegetale. • La crescita in LUNGHEZZA di fusti e di radici è presente in tutte le piante ed è definita CRESCITA PRIMARIA. Essa dipende dai meristemi situati ai due poli della pianta, apice vegetativo e apice radicale: cellule embrionali che continuano dividersi per mitosi. • Il corpo primario è la parte di pianta prodotta dai meristemi apicali. Moltissime piante, per esempio tutte le piante erbacee, hanno solo la struttura primaria perché l’ intero loro corpo è stato formato dai meristemi apicali. Gli alberi hanno anche una crescita secondaria ad opera dei meristemi secondari o cambi. La crescita in larghezza dipende dai meristemi secondari Meristema dell’apice vegetativo che permette la crescita in altezza della pianta Schema di una pianta con crescita secondaria, in sezione longitudinale e trasversale Localizzazione del meristema secondario (in nero): Cambio cribrovascolare Cambio Tessuti definitivi • • • • Tegumentali Parenchimatici Meccanici o di sostegno Conduttori o vascolari Tessuti Tegumentali Hanno funzione di protezione. I t. tegumentali rivestono le parti aeree e limitano traspirazione troppo elevata; sono costituiti da cellule a stretto contatto tra loro, senza spazi intercellulari, con pareti verso l’esterno impregnate da polimeri impermeabili quali cutina o la suberina. Il tessuto tegumentale delle parti non legnose della pianta, come fusti giovani, foglie, frutti, fiori, è l'epidermide, mentre le parti lignificate sono rivestite dal sughero. Anche la radice è rivestita, nelle parti non lignificate, da un'epidermide, il rizoderma, che ha una funzione completamente diversa: quella di assorbire acqua e sali minerali dal terreno: anche qui le cellule sono strettamente a contatto tra di loro ma hanno parete molto permeabile e alcune cellule possiedono delle lunghe estroflessioni, i peli radicali, che servono per aumentarne la superficie assorbente. Epidermide Riveste le foglie, i fusti erbacei, i fiori e i frutti; controlla il movimento dell'acqua tra interno ed esterno della pianta, la protegge contro altri organismi (es. funghi, batteri ed insetti) e contro agenti non biologici (es. vento). E' costituita da tre tipi di cellule: 1. Cellule ordinarie. Sono cellule vive, di forma variabile, strettamente a contatto tra loro, mancano di cloroplasti e sono trasparenti in modo da far arrivare la luce al parenchima clorofilliano sottostante. Le loro pareti esterne presentano cutina, molto impermeabile, e riflettente le radiazioni solari di protezione da funghi e batteri. Verso l’esterno si trovano anche delle cere (patina biancastra, la "pruina", di prugne e acini d'uva). 2. Stomi. "buchi" ad apertura regolabile, attraverso i quali avvengono la traspirazione e gli scambi di gas tra l'interno della pianta e l'ambiente esterno. Ogni stoma è costituito da due cellule di guardia, reniformi, che delimitano il poro stomatico. Quando le cellule di guardia assorbono acqua, si allungano e divaricano aprendo il poro stomatico, viceversa quando perdono acqua lo richiudono. Segnali quali la luce e la carenza di anidride carbonica nei tessuti fotosintetici danno il via ad un processo che si conclude con l'apertura dello stoma, mentre il buio, la carenza d'acqua, il calore troppo elevato ne determinano la chiusura. 3. Tricomi o peli. Comprendono tutte le strutture costituite da una o più cellule che si proiettano al di fuori del piano dell'epidermide; possono avere funzione protettiva (peli di rivestimento) oppure produrre sostanze di vario tipo: oli essenziali, sostanze urticanti, enzimi proteolitici, e così via (peli ghiandolari). Regno: Plantae Divisione: Magnoliophyta (Angiospermae) Classe: Liliopsida (Monocotyledones) Ordine: Alismatales Famiglia: Araceae Sottofamiglia: Aroideae Tribù: Arisareae Arisarum vulgare Targ. Tozz. Arisarum vulgare I peli o tricomi possono essere dei seguenti tipi : a) PROTETTORI Glabro, pubescente e tomentoso b) URTICANTI I peli urticanti cellule ghiandolari vive a maturità con SECRETO IRRITANTE (reazione istaminica). L’apice del pelo è impregnata di SILICE (SiO2) e quella basale di CARBONATO DI CALCIO (CaCO3). La mineralizzazione delle pareti cellulari rende il pelo rigido ma fragile, così penetra facilmente nella pelle e si spezza facendo uscire il suo veleno. ( Urtica dioica ) c) RADICALI sono unicellulari, con il nucleo in posizione distale e il citoplasma occupato da un grosso vacuolo. La parete cellulare è sottilissima e priva di cera in relazione alla loro funzione di assorbimento. Nelle plantule sopra raffigurate si vedeno la radice, il fusto, le foglie cotiledonari. Le plantule sulla sinistra non si sono ancora liberate dei tegumenti del seme, di colore scuro. In tutte le plantule sono evidenti i moltissimi peli radicali. Una forte riduzione del numero dei peli si osserva nelle piante con una buona SIMBIOSI MICORRIZICA. La MICORRIZA è una SIMBIOSI OBBLIGATA tra un fungo e le radici di una pianta (“mico”: fungo – “riza”: radice). TEGUMENTO SECONDARIO: SUGHERO Deriva dal meristema secondario che si ritrova nelle parti legnose dei fusti di Gimnosperme e Dicotiledoni legnose a livello della corteccia: cambio subero-fellodermico, o periderma, che produce - verso l'esterno il sughero, privo di spazi intercellulari, costituito da cellule rettangolari in sezione trasversale con parete cellulare suberificata, morte e contenenti aria a maturità. Il sughero quindi è un tessuto impermeabile ed è anche un isolante termico. - verso l'interno il felloderma, in quantità molto minore rispetto al sughero, costituito da cellule parenchimatiche Le pareti delle cellule del sughero sono impregnate di SUBERINA, una cera totalmente impermeabile, si tratta quindi di cellule morte a maturità. Per garantire comunque gli scambi gassosi ai tessuti vivi sottostanti, la disposizione molto regolare delle cellule del sughero è interrotta dove ci sono le LENTICELLE che infatti presentano ampi spazi tra le cellule per il passaggio dell’ aria. betulla Periderma: sostituisce l’epidermide nel corpo secondario di fusto e radice. E’ un tessuto formato dall’insieme di sughero, fellogeno , felloderma Sezione fusto di edera con lenticella RIzoderma il rizoderma è il tessuto di rivestimento della radice primaria, è monostratificato, con parete sottile, pectocellulosica nella zona assorbente e corrisponde in gran parte alla zona pilifera. Deriva dal meristema radicale, ed è formato da normali cellule appiattite provviste o meno di peli radicali. Al contrario dell’epidermide, il rizoderma ha pareti prive di cuticola e manca di stomi, quindi è del tutto permeabile ad acqua, sali minerali del terreno, e ai gas che sono convogliati verso i tessuti più profondi della radice. Non essendo esposto alla luce, le sue cellule sono prive di cloroplasti. Man mano che la radice si allunga, per opera del meristema apicale, nuovo rizoderma e nuovi peli si formano. Le cellule più giovani sono le più vicine alla cuffia radicale. Dopo la morte e la disgregazione (caduta) del rizoderma, lo strato di cellule corticali sottostanti suberifica e si trasforma in un tessuto tegumentale, sempre di origine primaria, l’esoderma. Esoderma Ha una funzione temporanea: ricopre la zona del rizoderma che va degenerandosi durante l’accrescimento delle radici. In particolare protegge la radice primaria nel passaggio a secondaria mentre si forma il sughero. E ’ presente nella parte più esterna del cilindro corticale. Funzione protettiva della radice primaria, con pareti cellulari ispessite nello strato più esterno per deposizione di suberina Endoderma E’ lo strato più interno del cilindro corticale adiacente alla parte più esterna del cilindro centrale (periciclo) Nelle radici forma un strato unicellulare senza spazi intercellulari. Le pareti hanno una fascia impregnata con materiale impermeabilizzante (suberina) che corre come un nastro lungo le pareti radiali e trasversali formando la banda di Caspary. In corrispondenza di questa fascia il plasmalemma aderisce fortemente alle pareti. Le cellule dell’endoderma esplicano la funzione protettiva controllando l’assorbimento minerale: obbligano l’acqua e i soluti assorbiti dalla radice a passare all’interno del loro citoplasma prima di raggiungere il cilindro centrale. Nel parenchima corticale acqua e soluti possono muoversi attraverso tre strade distinte: a) Apoplastica: spazi intercellulari, b) Simplastica: pareti cellulari, c) Transcellulare: citoplasma delle cellule. A livello delle cellule endodermiche, le bande del Caspary induco l’H2O e soluti ad attraversare la cellula (transcellulare). Avviene così il filtraggio dei soluti estranei. Le cellule dell’endoderma radicale che hanno smesso di funzionare, diventano lignificate con ispessimento a forma di U (nelle monocotiledoni)
Scaricare