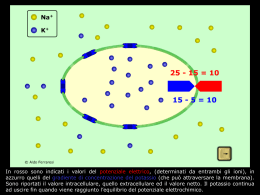
Tutte le cellule (non solo le cellule eccitabili) hanno un potenziale di riposo (resting): una carica elettrica attraverso la membrana plasmatica, con l’interno della cellula negativo rispetto all’esterno. Il valore del potenziale di riposo varia, ma nelle cellule eccitabili si aggira tra -90 e -70 mV. Il potenziale di membrana di un neurone a riposo misura circa -70 millivolt (mV), cioè il potenziale di un neurone all’interno della cellula è minore di circa 70 mV rispetto al liquido extracellulare. Il neurone allo stato di riposo è polarizzato. Il potenziale di riposo origina da: a) Ineguale distribuzione delle specie ioniche tra il liquido extracellulare ed intracellulare: INTRA (mM) EXTRA (mM) Na+ 12 145 K + 150 4,1 Ca2+ 10-7 (M) 3 Cl - 4 118 A- 146 1 CATIONI: ANIONI: b) Differente permeabilità della membrana ai diversi tipi di ioni: allo stato di “resting”, gli ioni potassio (K+) possono attraversare la membrana facilmente, mentre ioni cloro (Cl-) e sodio (Na+) presentano molte difficoltà. Le grosse molecole proteiche si comportano come ioni carichi negativamente (A-) e non attraversano la membrana. Due forze agiscono su una particella carica: La FORZA DI DIFFUSIONE, generata dal gradiente di concentrazione La FORZA ELETTRICA, generata dal gradiente elettrico La concomitanza di queste due forze spinge il flusso ionico all’interno o all’esterno della cellula. Cosa accade alla membrana quando è permeabile ad un solo ione? Nessuna differenza di potenziale Cosa accade alla membrana quando è permeabile ad un solo ione? Nessuna differenza di potenziale Cosa accade alla membrana quando è permeabile ad un solo ione? Cosa accade alla membrana quando è permeabile ad un solo ione? Cosa accade alla membrana quando è permeabile ad un solo ione? Si raggiunge l’equilibrio elettrochimico: La forza dovuta al gradiente di concentrazione eguaglia la forza dovuta al gradiente elettrico. FLUSSO NETTO NULLO. POTENZIALE DI EQUILIBRIO DELLO IONE PERMEANTE La relazione che esiste all’equilibrio tra gradiente chimico e gradiente elettrico fu stabilita dal chimico-fisico tedesco Walter Nernst: RT [X]E E x ln zF [ X] I Ex= Potenziale di equilibrio per lo ione X; R= costante dei gas T= temperatura assoluta (Kelvin) z= valenza dello ione F= costante di Faraday (96500 coulomb/grammo equivalente di carica) [X]i = Concentrazione intracellulare dello ione X [X]E= Concentrazione extracellulare dello ione X L’equazione di Nernst si ricava a partire dalla considerazione teorica: WE = WC (all’equilibrio) WE = zF Ex WC = RT ln [X]E /[X]I Quindi: zF Ex = RT ln [X]E /[X]I Da qui: Ex = RT/zF ln[X]E /[X]I Equazione di Goldman L’equazione di Goldman permette di descrivere in modo quantitativo il potenziale di membrana. In definitiva può essere considerata come una approssimazione dell’equazione di Nernst che tiene conto della permeabilità della membrana cellulare ai diversi ioni: RT Na+e PNa + K+e Pk + Cl-i PCl Vm = ln zF Na+e PNa + K+e PK + Cl-e PCl -PK ,PNa e PCl sono le costanti di permeabilità delle principali specie ioniche nei compartimenti intra ed extracellulari. EQUILIBRIO DI DONNAN Membrana impermeabile ad una sola delle specie ioniche presenti to t1 flusso dovuto al gradiente di concentrazione flusso dovuto al gradiente elettrico t2 Quando i flussi unidirezionali contrapposti saranno di uguale intensità per entrambi gli ioni, il flusso netto transmembranario delle specie diffusibili sarà nullo e si avrà: Vm = EK= ECl Applicando l’equazione di Nernst si ottiene: [ K ]1 [Cl ]2 [ K ]2 [Cl ]1 Quindi: Cl-1K+1=Cl-2K+2 Tale è l’equazione di Donnan dalla quale derivano 2 importanti conseguenze: 1) Ineguale concentrazione degli anioni e dei cationi diffusibili dal lato della membrana ove si trova la specie ionica non diffusibile. Per il principio di elettroneutralità delle soluzioni si avrà: K+1=Cl-1 (nel compartimento 1) K+2=Cl-2 + Pr-2 (nel compartimento 2) Quindi: K+2>Cl-2 2) La concentrazione totale degli ioni diffusibili è maggiore dal lato ove si trova la specie ionica non diffusibile. Dall’equazione di Donnan: Cl-1K+1=Cl-2K+2 Poiché: Allora: K+1=Cl-1 K+2>Cl-2 Cl- 2+K+2 >Cl- 1+K+1 Per “effetto Donnan” si produce a cavallo della membrana: a) Una differenza di potenziale stabile nel tempo; b) Un aumento della pressione osmotica del liquido ove sono presenti i proteinati. mantenimento turgore cellulare E’ possibile considerare la membrana come un conduttore ohmico attraverso cui fluiscono correnti ioniche specifiche. Dato lo ione X,esso produrrà una corrente avente intensità: IX = (Vm - EX) gX Dove gX è la conduttanza della membrana nei riguardi dello ione X. Assumendo che in una cellula eccitabile: Vm= -70 mV Poiché ECl è quasi uguale a Vm, ENa= +60 mV allora si può accettare che il EK= -84 mV contributo degli ioni cloruro al ECl= -72 mV potenziale di membrana sia nullo. Alla genesi del potenziale di membrana partecipano, dunque, solo sodio e potassio che sostengono due correnti ioniche transmembranarie: Cl- Pr- Na+ a) IK diretta dall’interno all’esterno della cellula e generata dal gradiente di concentrazione. b) INa diretta dall’esterno all’interno della cellula e dovuta sia al gradiente di concentrazione,sia al gradiente elettrico. All’equilibrio elettrico, che caratterizza il potenziale di riposo,le due correnti ioniche saranno di uguale intensità: IK =-INa Per cui la sommatoria delle correnti: INa + Ik + Icl = 0 Da questa deduzione si ricava la legge della conduttanza: Vm= [(gNaENa+gKEk+gClECl)+(INa+IK+ICl)]/(gNa+gK+gCl) Tale legge permette di predire il potenziale di riposo della membrana IL RUOLO DELLA POMPA Na+/K+ NELLA GENESI E NEL MENTENIMENTO DEL POTENZIALE DI MEMBRANA La pompa Na+/K+ ripristina il gradiente ionico pompando attivamente (idrolisi ATP) 3Na+/2K+ rispettivamente all’esterno e all’interno della cellula. La cellula raggiunge lo stato stazionario quando il Vm sarà tale che i due flussi attivi creati dalla pompa sono di uguale intensità ai flussi passivi dei due ioni permeanti attraverso gli appositi canali(passivi).
Scaricare