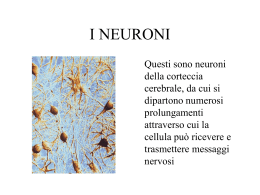
Meccanismo ionico dei potenziali postsinaptici EPSP •Aumento di conduttanza a Na+ e K+ IPSP •Aumento di conduttanza a K+ oppure •Aumento di conduttanza a Cl - Gli EPSP e IPSP vengono condotti elettrotonicamente. Il potenziale di azione si genera nel cono di emergenza (monticolo assonico - segmento iniziale) Sommazione dei potenziali postsinaptici Input 1 Input 2 Sommazione spaziale Input1 Input2 Input 1 + 2 Sommazione temporale Input1 Input1+ Input1 Inibizione presinaptica Viene inibito il rilascio di neurotrasmettitore da parte di una sinapsi eccitatorio Il neurone postsinaptico è “inibito” in quanto non viene depolarizzato Rigenerativo Canali voltaggio dipendenti Fenomeno tutto o nulla Refrattarietà Aumento P Na e poi K Grande ampiezza (100 mV) Inversione della polarità Propagazione attiva Non rigenerativo Canali ligando dipendenti Fenomeno graduale Sommazione Aumento simultaneo P Na e K Piccola ampiezza ( 10 mV) Depolarizzazione Propagazione locale Neurotrasmettitori •A basso peso molecolare •Peptidi neuroattivi I neurotrasmettitori a basso peso molecolare (“classici”) sono sintetizzati nel terminale assonico I peptidi neuroattivi sono sintetizzati nel soma (reticolo endoplasmatico). Vengono trasportati entro vescicole, tramite trasporto assonico rapido, verso il terminale assonico Neurotrasmettitori a basso peso molecolare Catecolamine ACETILCOLINA •Motoneuroni midollo spinale •Motoneuroni della corteccia motoria •Gangli della base •SNA AMINE BIOGENE Catecolamine. sintetizzate a partire da tirosina. •La noradrenalina è il neurotrasmettitore dei neuroni ortosimpatici postgangliari •Le cellule della midollare surrenale aggiungono un gruppo metilico alla noradrenalina e formano adrenalina •La dopamina è il neurotrasmettitore dei neuroni della substantia nigra che proiettano al corpo striato (nel morbo di Parkinson si verifica degenerazione dei neuroni dopaminergici AMINOACIDI La glicina è un neurotrasmettitore inbitorio di interneuroni spinali L’acido g-aminobutirrico (GABA) è un neurotrasmettitore inbitorio sintetizzato da glutammato (decarbossilasi specifica presente solo in alcuni neuroni). E’ molto diffuso nel cervello La glicina e il GABA iperpolarizzano la cellula in quanto i loro recettori sono canali per Cl-. Gli anestetici generali prolungano l’effetto inibitorio Glutammato e aspartato sono aminoacidi dicarbossilici. Hanno potente effetto eccitatorio. Sono i trasmettitori eccitatori più diffusi MONOSSIDO DI AZOTO •Neuroni del sistema nervoso enterico •SNC E’ un neurotrasmettitore che non viene immagazzinato in vescicole. E’ altamente diffusibile. Deriva dalla ossidazione di arginina o citrullina ad opera della NO sintasi (stimolata da un aumento di Ca++ nel citosol) Peptidi Neuroattivi Sono stati identificati circa 25 neuropeptidi Sono formati da una catena di 2-40 aminoacidi In genere la loro azione è pià prolungata rispetto ai neurotrasmettotori “classici” Alcuni neuropeptidi sono neurotrasmettitori (modificano la conduttanza ionica del neurone postsinaptico) Alcuni neuropeptidi sono neuromodulatori (modulano la trasmissione sinaptica, ad esempio facendo variare la quantità di neurotrasmettitore liberato) Possono essere co-localizzati con i neurotrasmettitori “classici” nello stesso terminale Le vescicole che li contegono non sono ancorate a zone attive. Il loro rilascio richiede l’ingresso di una elevata quantità di calcio Trasmettitore dei neuroni primari che mediano la nocicezione Oppiacei - Sostanze che derivano dall’oppio del papavero. Hanno potente effetto analgesico. Si legano a specifici recettori per gli oppiacei. Il legame è bloccato dal naloxone Peptidi oppiodi - Sono sostanze endogene la cui azione è bloccata dal naloxone. •Encefaline •Endorfine Recettori per i neurotrasmettitori Recettori ionotropi - Sono canali ionici regolati da ligandi . Mediano fenomeni postsinaptici che si esauriscono in pochi msec. Recettori metabotropici - Non sono canali ionici. Mediano la prima fase del processo di trasduzione del segnale che modifica la funzione del canale ionico. Mediano fenomeni postsinaptici che possono durare diversi msec-minuti. L’apertura/chiusura del canale dipende da un secondo messaggero Esempi di recettori ionotropi: il recettore nicotinico per Ach, i recettori ionotropi per il glutammato, il recettore per il GABA (tipo GABAA) Esempi di recettori metabotropici: i recettori per il GABA (tipo GABAB) e i recettori metabotropi per il glutammato Secondi messaggeri a classi di proteinechinasi Il legame di un agonista con il suo recettore provoca l’aumento nel citoplasma della concentrazione di un secondo messaggero che aumenta l’attività di una proteina chinasi La proteina chinasi provoca la fosforilazione di proteine. La fosforilazione può aumentarne o diminuirne l’attività. •I canali ionici possono essere direttamente regolati dalla proteine G (senza partecipazione di secondi messaggeri) •L’apertura chiusura di un canale può dipendere dai livelli di secondo messaggero Recettori muscarinici M2 per Ach del cuore e di alcuni neuroni Attivazione di una proteina Gi Interazione del dimero g con un canale K+ -> Apertura del canale Aumento uscita K+ -> Iperpolarizzazione L’Ach diminuisce con questo meccanismo la frequenza cardiaca Le proteine G sono intermediari tra i recettori di membrana e i processi intracellulari Gs Gi Esempio: Regolazione dell’adenilato ciclasi Adenilato ciclasi Gs Gi Stimolazione Inibizione Esempio: Regolazione della Fosfolipasi C (PLC) Recettori per adrenalina e noradrenalina (1 2) Tipo 1 2 1 2 (1 2) Risposta NA>A>ISO NA>A>ISO ISO>A=NA ISO>A>NA Meccanismo di azione Ca++ AC AC AC AC, adenilato ciclasi; A, Adrenalina; NA, noradrenalina; ISO, isoproterenolo
Scaricare