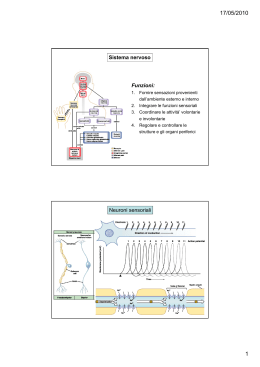
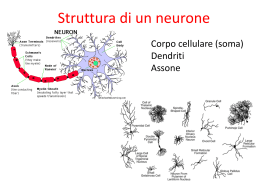
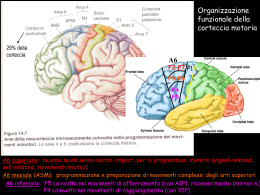
10. Sistemi sensoriali Per sensazione intendiamo un processo elementare di percezione e ricezione di un singolo stimolo, mentre la percezione è un'analisi molto complessa di stimoli. Ogni esperienza sensoriale è determinata da tre componenti: stimolo, codice neuronale e rappresentazione mentale. Per Comte la mente è una tabula rasa, quindi la percezione era un processo passivo di analisi delle singole componenti sensoriali. La posizione della Gestalt dà un ruolo attivo alla mente: l'oggetto stimolo viene codificato nel sistema sensoriale, ma la codifica diventa un processo attivo. Un esempio di regola innata sono le sfere. Si vuole dimostrare è che quando la superficie viene illuminata sotto il tipo di informazione percettiva che abbiamo è quella di qualcosa che risulta profondo, al contrario il pattern reciproco, in cui la parte illuminata è sopra, rappresenta una sfera, perchè ci dà la sensazione di rilievo. Ruotando di 90° gli stimoli, non abbiamo più a stessa percezione. Il sistema visivo si è sviluppato avendo come fonte di illuminazione il sole, che illumina dall'altro, quindi tende a considerare le superfici come ombre e luci in posizioni differenti in base a ciò che succede quando la luce viene da sopra. Un esempio di regola acquisita è il parametro interno culturale: l'immagine mostra una famiglia africana, mentre il campione africano ha descritto l'immagine come un gruppo di persone che si trova sotto un albero, la signora tiene qualcosa sulla testa e si afferma che sta avvenendo una tratta di schiavi. Quindi l'esperienza sensoriale è un processo attraverso il quale il sistema sensoriale percepisce lo stimolo fisico e ne estrae determinate caratteristiche che trasforma in esperienze sensoriali o percettive (colori, suoni, odori) che sono costrutti mentali non esistenti nel mondo esterno, fortemente influenzati da regole innate o acquisite (culturali). Lo stimolo proviene dall'ambiente esterno in generale, va ad agire su un determinato elemento del nostro organismo, che è un recettore. Il recettore, che può essere una cellula specializzata indipendente o una sua porzione, trasduce l'informazione sensoriale e lo invia a un neurone sensitivo primario situato a livello del SNP. Il neurone sensitivo primario invia l'informazione dalla periferia a livello centrale e inviano l'informazione ulteriormente a strutture sottocorticali come talamo e corteccia. A questo punto ottengo la consapevolezza cosciente della stimolazione sensoriale. La via che serve per un controllo motorio è quella che parte dallo stimolo, attivando un recettore, arriva al midollo spinale e causa una risposta riflessa. Un'altra (di controllo sul livello di attivazione dell'organismo) è quella che, dal recettore,. Una terza raggiunge la consapevolezza cosciente attraverso la corteccia cerebrale. I recettori sono di vari tipi. Una prima classificazione riguarda il meccanismo di trasduzione. Da questo punto di vista abbiamo i meccanocettori (attivati a seguito di una distorsione fisica del recettori), i chemocettori (riescono a rilevare la presenza di sostanze chimiche nell'ambiente, hanno una proteina che si lega alla sostanza e così procedono all'attivazione), i termocettori (rilevano la temperatura), fotocettori (rilevano onde elettromagnetiche). Abbiamo una classificazione sulla base della posizione e in questo senso si parla di esterocettori (trasmettono informazioni dalla superficie del corpo), gli enterocettori (trasmettono le informazioni dalle viscere), i propriocettori (trasmettono le informazioni dai muscoli e ossa). Una terza classificazione riguarda la morfologia: parliamo di recettori di I, II e III ordine. Quelli di I ordine sono quelli in cui il recettore è una parte specializzata del neurone sensitivo primario. I recettori di III ordine sono esclusivamente i fotorecettori, quindi solo quelli della retina. Questo non contatta direttamente il neurone sensitivo primario, ma tra i due elementi vi è un interneurone. Le caratteristiche dello stimolo sono essenzialmente quattro: modalità, intensità, durata e localizzazione. Per modalità si intendono i cinque sensi principali; a queste cinque bisogna aggiungerne altre. La modalità tattile comprende anche la nociocezione (percezione del dolore), la termocezione (percezione della temperatura) e la propriocezione (percezione della propria posizione nello spazio). Si parla anche di submodalità: ciascuna delle modalità principali hanno submodalità, per esempio il gusto ha tre submodalità: amaro, dolce, salato. Vista: colore, forma, movimento. Secondo la linea risevata, modalità e submodalità dipendendono dalla via sensoriale interessata: indipendentemente da come si attiva una modalità, la sensazione finale è quella specifica di ogni modalità. Viceverva, quando si parla di stimolo adeguato, si parla della categoria di stimoli che riesce ad attivare una determinata modalità con minor contenuto energetico. L'intensità dello stimolo è il contenuto di energia dello stesso; quando parliamo di sostanze chimiche, l'intensità dello stimolo sarà la concentrazione della sostanza. L'intensità è codificata in termini diversi in base al tipo di stimolo. Accanto al concetto di intensità si deve considerare il concetto di soglia. La soglia, in generale, è l'intensità dello stimolo oltre alla quale riesco a ottenere una sensazione. Esistono soglie diverse: soglia del recettore, soglia di percezione e soglia differenziale. La soglia del recettore è l'intensità minima dello stimolo necessaria per attivare un recettore. La soglia di percezione è . la percezione dipenderà poi dalla detettabilità e dalla sensibilità. La prima dipende dal sistema sensoriale, la seconda dalla nostra attenzione allo stimolo. La soglia differenziale riguarda la capacità di percepire le più piccole differenze tra due stimoli. La durata è la permanenza nel tempo di uno stimolo, permanenza che riesco a descriminare sulla base della codifica dello stimolo stesso. La durata di uno stimolo è associata al concetto di adattamento: vi sono tipi di stimoli che attivano recettori a seconda del loro adattamento La localizzazione permette la discriminazione spaziale, massima sulla punta delle dita. Nella condizione di riposo non ho nessuno stimolo e nessun potenziale d'azione, ma il neurone è comunque attivo. Lo stimolo, se soprasoglia, riesce ad attivare il recettore, producendo un potenziale di recettore, di ampiezza diversa a seconda dell'intensità dello stimolo. Il potenziale di recettore genera un potenziale postsinaptico. Se quest'ultimo è soprasoglia genera un potenziale d'azione. C'è una relazione (legge di Stevens) tra intensità dello stimolo e intensità della sensazione percepita. I = k (S – S0)n k = costante I = intensità percepita S0 = stimolo soglia S = stimolo n dipende dalla modalità sensoriale Abbiamo recettori che non adattano o che hanno un adattamento lento, rapido e rapidissimo. I recettori senza adattamento rispondono allo stesso modo per tutta la durata dello stimolo. Quelli con adattamento lento rispondono principalmente all'inizio Quelli con adattamento rapido rispondono principalmente all'inizio e alla fine. Il rpocesso di adattamento corrisponde a un decremento della sensazione al persistere dello stimolo. Per unità recettrice si intende il recettore con tutti i neuroni sensoriali a cui invia informazioni. Abbiamo sistemi divergenti e sistemi convergenti. Nei primi abbiamo lo stesso recettore che invia l'informazione a più di un neurone sensitivo primario; al contrario, nei sistemi convergenti abbiamo lo stesso neurone sensitivo primario che riceve l'informazione da più recettori. Un sistema convergente permette un aumento della sensibilità, mentre il sistema divergente permette una maggiore acuità e discriminazione, perchè lo stesso recettore attiva più di un neurone sensitivo primario. Nell'elaborazione corticale ci sono due meccanismi: uno di tipo gerarchico, in cui uno stesso stimolo viene scomposto nei diversi elementi che lo costituiscono. L'analisi parallela sostiene che lo stimolo viene scomposto nelle sue submodalità, come forma, colore e movimento, processate parallelamente per poi ricostruire la conformazione unitaria. 11. Il sistema somatoestetico La modalità tatto comprende altre modalità. Oltre al tatto propriamente detto c'è la propriocezione, la nociocezione e la termocezione. Quando parliamo di sistema somatoestesico parliamo di un sistema che porta informazioni in tutto il corpo. Gli esterocettori portano informazioni dalla superficie del corpo e sono distribuiti ovunque nel corpo. I propriocettori sono i recettori che portano informazioni da ossa, muscoli e articolazioni e servono a dare informazione non solo sulla posizione delle parti del corpo, ma anche sulla posizione dell'individuo nello spazio. Gli enterocettori sono stimolati e trasmettono informazioni dai visceri. Una prima differenza del sistema somatosensoriale è che i recettori non sono legati a un organo specifico, ma sono distribuiti ovunque nella superficie corporea. Un'altra caratteristica è che i recettori che abbiamo visto del sistema somatosensoriale sono di I ordine, eccetto quelli del volto. Di conseguenza la cellula recettrice non è un elemento indipendente ma è una porzione specializzata del neurone sensitivo primario, che in questo caso si trova nei gangli delle radici dorsali. Una differenza sostanziale è tra i recettori della fibra pelosa e i recettori della fibra glabra; quelli della cute pelosa sono dei meccanocettori, avvolti attorno alla superficie del pelo e si attivano quando c'è uno stiramento del pelo; i recettori della cute glabra sono essenzialmente localizzati nelle porzioni di corpo non avvolte dai peli, come la mano e le dita. I dischi di Merkel sono localizzati essenzialmente in corrispondenza delle impronte digitali. Occorre sapere che rappresentano il 25% circa dei recettori della mano e sono recettori ad adattamento lento. È stato dimostrato che sono recettori particolarmente sensibili a punti, spessori e curvature e servono proprio alla percezione della forma e delle strutture superficiali degli oggetti. La stereognosi è la capacità della mano di riconoscere un oggetto, mediata dalle caratteristiche di questi recettori. I corpuscoli di Meisner sono subito sotto la cresta; in questi colpisce la forma allungata che possiedono. Tutte queste strutture nonostante siano specializzate, non sono elementi autonomi, ma sono parti del neurone sensitivo primario. Il corpuscolo rappresenta il 40% dei recettori situati nella mano ed è un recettore ad adattamento rapido; è costituito da una capsula che contiene lamine di cellule di Shwann. Sono importanti per la percezione del movimento e il controllo della presa. I corpuscoli di Pacini sono nella porzione più profonda dell'epidermide, la loro formologia è costituita da una serie di strati concentrici di lamelle che circondano la singola fibra apparente. Sono recettori ad adattamento rapido e servono a percepire gli eventi lontani attraverso la trasmissione di vibrazioni e sono essenziali per poter utilizzare gli strumenti. L'ultimo tipo di recettore è il corpuscolo di Ruffini che rappresenta il 20% e sono ad adattamento lento; sono strutture allungate, localizzate nella profondità della cute ma ce ne sono anche nei legamenti e nei tendini. I corpuscli sono sensibili allo stiramento delle dita e della cute e rilevano ogni tipo di stiramento che facciamo con la mano. Vi sono anche terminazioni nervose libere, che mediano le informazioni dolorifiche termiche. L'elemento centrale del sistema tattile propriocettivo e somatosensoriale in generale, è il neurone sensitivo primario, che ha un caratteristico assone a T, in cui un'estemità termine nelle terminazioni nervose libere nel caso del sistema dolorifico e l'altra va a connettersi nel neurone sensitivo secondario che poi si connetterà al talamo e corteccia. Alla corteccia si ha una sensazione soggettiva e cosciente. I corpi cellulari di questi neuroni sensitivi primari del sistema somatosensoriale sono nei gangli delle radici dorsali e quindi vi è una terminazione che va nell'epidermide e l'altra porzione entra a livello del midollo spinale proprio dalle radici dorsali. Quando parliamo di acuità tattile parliamo della capacità di discriminare due stimoli attraverso il tatto. Questa acuità tattile è una funzione sia delle dimensioni dei campi recettivi sia della densità dei recettori; nel caso della mano, questa ha recettori con campi recettivi piccoli sulle dita, viceversa aumenta nelle altre parti della mano come nel palmo. Il campo recettivo è la porzione della periferia sensoriale la cui stimolazione porta all'attivazione di un determinato recettore. La capacità di acuità tattile si può misurare prendendo le due punte del compasso e provando a diminuire la distanza fino a quando non si riesce più a distinguere le due stimolazioni differenti. Nelle zone che hanno campi recettivi più piccoli e maggiore densità di recettore l'acuità tattile si traduce in una distanza minima tra i punti, quindi l'acuità è molto elevata (2 mm sulla punta delle dita, 4 mm sul dito, 8 mm sul palmo della mano). I recettori che hanno campi recettivi piccoli (che permettono una discriminazione più fine dello stimolo) e adattamento lento (sono attivi per tutto il tempo di discriminazione dell'oggetto) servono alla discriminazione spaziale dell'oggetto. Viceversa, recettori con campi recettivi di dimensioni maggiori con rapido o rapidissimo adattamento consentono una discriminazione temporale dello stimolo. Se l'acuità tattile diminuisce, le zone avranno meno recettori e saranno di dimensioni maggiori. Il sistema tattile è chiamato sistema delle colonne dorsali: ha come elemento centrale le cellule con corpi cellulari nei gangli delle radici dorsali, che sono i neuroni sensitivi primari. Questi hanno una porzione del loro assone a T che termina nelle terminazioni del recettore (linea rossa) e questo ci fa comprendere che ci sono fibre afferenti che possono essere anche molto lunghe, di mezzo metro di lunghezza. L'altra porzione del corpo cellulare nel ganglio delle radici dorsali entra nel midollo spinale. Le fibre che inviano informazioni relative agli arti inferiori sono situate nella porzione mediale e viaggiano in fasci ben definiti e circoscritti (fascicolo gracile). Le fibre che inviano informazioni relative agli arti superiori, al tronco e al collo si trovano in fasci posti lateralmente e conosciuti come fascicolo cuneato. Le fibre di questi fasci terminano in distinte suddivisioni dei nuclei delle colonne dorsali: il nucleo gracile e cuneato. L'informazione relativa ai diversi tipi di recettori viene sempre elaborata in modo separato; ogni recettore ha un suo canale parallelo. Ci saranno fibre specifiche che prendono le informazioni e tutto questo ha un canale saparato anche da un punto di vista anatomico fino alla corteccia. Ci sono diversi processi di inibizione: una è l'inibizione laterale, che consiste nel fatto che ho una singola fibra afferente che contatta il neurone sensoriale, ma la stessa fibra ha dei collegamenti con A, B e C rappresentano neuroni in aree limitrofe della superficie laterale. Questi ricevono informazioni dalla stessa zona del campo recettivo ma da aree diverse. Il neurone B aumentano la frequenza di scarica, mentre i neuroni A e C diminuiscono la frequenza a causa dell'interneuone inibitorio. La fibra afferente va ad attivare sia il neurone B sia gli interneuroni; gli interneuroni inibitori sono quelli che non consentono ad A e C di scaricare l'impulso. Ha funzionamento per contrasto. L'inibizione laterale permette la discriminazione tra due punti. I neuroni inibitori sopprimono l'attivitazione dei neuroni disposti tra i due punti attivati, rendendo più nette le zone attivate. 3a, 3b, 1 e 2 sono sottoaree della corteccia S1 (corteccia somatosensoriale) sitata nell'area parietale. Se ci sono lesioni a livello di S1, le aree di S2 (corteccia di ordine superiore) sono difatto silenti, perchè riceve da S1. Il talamo non proietta S2, ma proietta solo 3a e b. L'informazione, dalle aree 3a e 3b, viene trasmessa alle aree 1 e 2 e infine l'informazione di tutte queste aree converge in S2. È un esempio di analisi gerarchica. Importanti sono le frecce blu che collegano S1 ed S2 alla corteccia motoria e parietale posteriore (area di associazione e integrazione di stimoli). All'interno delle sottoaree sono elaborate informazioni diverse delle caratteristiche dell'oggetto (forma, dimensione, struttura) parallelamente. Nei primani non umani, lesioni selettive dell'area 3b portano a un grave deficit della sensibilità tattile mediata dai meccanocettori cutanei. Le lesioni delle aree 1 e 2 portano a deficit pariziale e inabilità nell'uso delle informazioni tattili per la discriminazione... La rappresentazione somatotopica è stata studiata da Marshall, il primo a effettuare le registrazioni nella corteccia del gatto dopo la stimolazione delle aree della cute. In seguito Kaas e Merzeniche hanno scoperto come le rappresentazioni sono di varia natura e sono contenute in diverse aree di S1, quindi non si parla solo di mappa somatotopica ma di sottomappe. Le mappe non sono statiche, ma plastiche nel tempo; variano in seguito a sviluppo, lesione ed esperienza. Arto fantasma in seguito a lesione: per un certo periodo si avrà la sensazione della sua presenza nonostante questo non ci sia (dovuta a riarrangiamento corticale). L'informazione arriva separata fino alla corteccia: questa è formata da una serie di moduli che processa una determinata parte di informazione. L'organizzazione è colonnare ed è un ulteriore esempio di processamento parallelo. 12. Sistema somatoestetico 2 Nel freddo paradosso la sensazione del freddo precede quella del bruciore. Le fibre δ (delta) sono mielinizzate (infatti sono più veloci), mentre quelle C sono amieliniche. I recettori per il freddo ha fibre mielizzate, quindi nel freddo paradosso (temperature maggiori a 45°), sentirò prima la sensazione del freddo, trasmetta dalle fibre mieliniche, poichè più veloci. All'aumentare delle temperature, aumenterà la frequenza di scarica. Nel caso di stimoli termici dolorosi l'intensità dello stimolo è legata all'aumento della temperatura. Il dolore non corrisponde alla nocicezione. La nocicezione comprende diversi tipi di sensazione tra cui il dolore. Per nocicezione si intendono una serie di informaizoni sensoriali che dichiarano il danno potenziale o il periclo di un danno tissutale in atto. La sensazione di nausea o mancamento è mediata dai nociocettori che non trasmetterò però l'informazione di una sensazione dolorifica. Il sistema responsabile di queste sensazione è il sistema nocicettivo. Questo è correlato alle nostre risposte di pericolo e allerta: ci sono tre tipologie di risposte di fronte a un pericolo: fuga, attacco e breazing. Nocicezione e dolore quindi non sono assolutamente la stessa cosa. Ci sono tre tipi di nocicettori: termici, meccanici e polimodali. I primi rispondono a stimoli termici (variazioni di temperatura) e possono rispondere a stimoli superiori a 45° e inferiori a 10°. Saranno attivi i call sport per il freddo paradosso. I meccanici rispondono a stimoli meccanici e i polimodali rispondono a stimoli termini, meccanici o chimici. Trasmettono il dolore sordo. Il dolore può essere descritto in modi diversi, sia perchè la soglia soggettiva è diversa da individuo a individuo. Tendenzialmente si distingue in tre categorie: acuto, persistente, cronico. Il dolore acuto è la sensazione di dolore comune. Il dolore persistente è l'esperienza di dolore di cui soffrono persone con patologie importanti come la sclerosi o l'AIDS, in cui è esperienza quotidiana, perchè tutte le vie di trasmissione nociocettiva sono compromesse. Il dolore cronico è una sensazione di dolore che deriva da una modifica plastica a livello corticale. Il dolore acuto si può dividere in dolore primario e secondario. I due tipi di dolore sono mediati da fibre di tipo diverso. Il dolore acuto primario è dominato da fibre gamma. Inibendo la trasmissione dei due tipi di fibre, la sensazione soggettivo è quella di provare solo un tipo o l'altro del dolore. Bloccando le fibre di tipo gamma l'esperienza soggettiva è quella di dolore secondario, di tipo sordo. Viceversa, bloccando le fibre C, vi è solo la prima sensazione immediata di dolore primario e poi cessa immediatamente e non si ha continuità dell'esperienza dolorifica mediata dalle altre fibre. La zona che rileva il dolore è più ampia rispetto a quella dove avviene il dolore. La sensazione soggettiva di dolore in zone più ampie è data dai policettori. L'esperienza è data solo dall'area A, ciò è meno conservativo per l'organismo in sè. Gli estremi sono tradotti dai concetti di iperalgesia e allodinia. L'iperalgesia è una reazione eccessiva agli stimoli dolorifici, quindi anche zone normali vengono coinvolte nel processo dell'infiammazione. L'allodinia è un dolore provocato da stimoli normalmente innocui. Entrambe però fanno parte del range delle 'patologie' del sistema nociocettivo proprio perchè riguardano il funzionamento aberrante dei meccanismi di segnalazione del dolore. Le terminazioni nervose libere provengono dal neurone primario. L'altra parte dell'assone entra tramite la radice dorsale ma va subito nella sostanza grigia del midollo spinale, a livello delle corna dorsali del midollo spinale. Il neurone primario contatta i secondari. Il primo tipo di connessione avviene proprio a livello delle corna. Le afferenze inviate da metà del corpo hanno subito una degussazione nel midollo spinale. I neuroni di II ordine risalgono seguendo un sistema di colonne anterolaterali, perchè sono fasci di fibre che attraversano la sostanza bianca e quindi attraversano le colonne e si dicono antero-laterali perchè la loro conformazione anatomica prevede una modificazione verso il piano antero-laterale. Il fascio neospinotalamico è essenziale ricordare che ha la sua stazione talamica nel complesso ventrovasale del talamo (III ordine) e da qui invia le informazioni finali. Gli altri tipi di fasci si comportano in modo analogo, salvo che la loro stazione sono i nuclei. Il paleo-spinotalamico è il fascio più antico, mentre quello neo-spinotalamico si è evoluto nelle specie recenti e nell'uomo. Più precisamente: Fascio neo-spinotalamico: va al talamo ventrobasale, che porta le informazione dolorifiche alla corteccia parietale somoestesica (SI): percezione dello stimolo dolorifico e sua localizzazione, anche se non c'è una mappa. Fascio paleo-spinotalamico: va al talamo (nuclei intralaminari), che proietta in maniera diffusa alle cortecce (arousal). Fascio spinoreticolare: va alla sostanza reticolare, che proietta a talamo (e quindi a cortecce) e ipotalamo (arousal, riflessi vegetativi). Fascio spinomesencefalico: va al grigio periacqueductale e da qui al talamo e poi alle cortecce, incluse la corteccia insulare e la corteccia cingolata anteriore, responsabili delle componenti affettive. Ha una connotazione negativa e riguarda le reazioni emotive ad uno stimolo dolorifico. Ci sono indizi sull'esistenza di una mappa del dolore, ma non è stata ancora identificata. La stimolazione con degli elettrodi i neuroni della sostanza grigia produce analgesia, che attiva la via centrale del dolore e si inibisce l'azione dolorifica. Il soggetto non comunica dolore. Invece di stimolare queste aree, si può somministrare morfina ottendendo lo stesso risultato (assenza di dolore). Viceversa, se si applica un aloxone, sostanza chimica che agisce sui recettori della morfina, l'effetto di quest'ultima scompare e si prova dolore. Tutte queste prove sperimentali hanno dimostrato che esistono all'interno delle vie discendenti esistono neuroni che esercitatono un'azione inibitoria del dolore e sono simili agli oppioidi (per questo chiamati...). È stato studiato il livello di contatto primario tra il neurone sensitivo primario con quello secondario per capire se anche a questo livello fossero presenti fattori di inibizione. È stato dimostrato come il neurone del corno dorsale ha dei recettori per gli oppioidi: delta, mu e K. Il principale è il recettore mu, che si lega agli oppioidi endogeni. 12. Sistema visivo Attaverso gli occhi vediamo qualsiasi stimolo visivo come un'immagine unitaria e tridimensionale. Ciò che però dobbiamo tenere in considerazione è che sulla retina l'immagine si forma con tre caratteristiche principale. L'immagine è, innanzitutto, doppia, capovolta e bidimensionale. Un'altra caratteristica del sistema visivo rispetto agli altri è il fatto che lo stimolo non è una proprietà intrinseca dell'oggetto. Quando parliamo di stimoli visivi non ci riferiamo a oggetti che hanno delle caratteristiche specifiche, ma lo stimolo vero e proprio è un segnale indiretto, emesso da una sorgente luminosa e riflesso dall'oggetto in questione. Lo stimolo sono i fotoni, le lunghezze d'onda, emesse da una sorgente luminosa e che ciascun oggetto riesce ad assorbire e riflettere. Ciò avviene a differenza degli odori, dei suoni o del gusto, che hanno proprietà intrinseche. Lo stimolo vero e proprio sono le onde elettromagnetiche, che si distribuiscono su un'ampia gamma di lunghezze d'onda. Tuttavia quelle relative allo spettro visibile sono comprese da 400 e 760 nanometri circa. Tra tutte le onde del visibile, quindi, i nostri occhi riescono a processare soltanto una parte. Le proprietà di un'onda elettromagnetica sono quelle di rifrazione, riflessione e assorbimento. In fisica, per quanto riguarda il processo di riflessione, sono stati studiati alcuni principi, due essenziali. Secondo il primo principio il raggio incidente, il raggio riflesso e la normale giacciono sullo stesso piano. Inoltre l'angolo di incidenza e l'angolo di riflessione sono esattamente uguali. Questo principio spiega come avviene la riflessione delle onde. Viceversa, quando parliamo di rifrazione, uno dei concetti centrali nell'organizzazione del sistema visivo, si parla di un processo secondo il quale un'onda viene deviata quando passa da un mezzo all'altro e in questo passaggio varia la velocità di propagazione. L'assorbimento si realizza quando abbiamo una porzione di materia che viene a contatto con la radiazione elettromagnetica e riesce ad assorbire l'energia della radiazione stessa. Tale assorbimento però dipende dalle proprietà della materia in sè e dalla lunghezza d'onda della radiazione. Vediamo com'è fatta la porzione periferica del sistema sensoriale visivo. Partiamo da una considerazione secondo cui l'occhio è sostanzialmente formato da due componenti. Vediamo grazie all'organo recettivo vero e proprio, la retina e a un sistema di lenti, detto sistema diottrico, il cui elemento principale è il cristallino. Questo sistema di lenti, che ha un livello di consistenza superficiale simile al vetro, serve per mettere a fuoco ogni punto sulla retina. Strutturalmente, a in prossimità del sistema diottrico, ci sono una serie di muscoli interni che servono a regolare il diametro della pupilla e l'accumulazione del cristallino. Un'altra funzione molto importante del cristallino è il fatto che filtra anche le radiazione non visibili. Tra le altre componenti abbiamo l'iride, che è la parte colorata dell'occhio. L'iride si può vedere attraverso la cornea, rivestimento protettivo che prosegue in tutto l'occhio, ma diventa da sclera, porzione di tessuto bianco fibroso. Lo spazio compreso tra la porzione posteriore del cristallino e la parte finale dell'osso bulbo-oculare è pieno di una sostanza detto umore vitreo, sostanza gelatinosa che occupa quasi l'80% del volume dell'occhio e contiene cellule che hanno un'attività fagocitaria, cioè vanno a rimuovere elementi del sangue o impurità che potrebbero interferire con la trasmissione della la luce. La superficie della retina è caratterizzata dalla presenza di una vascolarizzazione massicia, cioè ci sono sia vene che arterie, disposte a ventaglio, derivanti tutte dalla vena ottalmica, che entra nell'occhio attraverso il disco ottico. Quel punto è essenziale non solo perchè è il punto di ingresso di vene e arterie, ma anche perchè è il punto di uscita delle informazioni visive che poi saranno trasmette al nervo ottico e alle altre stazioni successive. La retina ha delle porzioni assolutamente specifiche. Una porzione specifica è la macula lutea, qui si trova la più alta densità di recettori e quindi si ha la massima acuità visiva, cioè la capacità di vedere i dettagli visivi con la più alta risoluzione possibile. Il centro della macula lutea è la fobea, che contiene un pigmento protettivo che serve a proteggere la retina dalle radiazioni ultraviolette, che potrebbero essere dannosi per i recettori retinici. Sia la cornea che il cristallino ha la proprietà di rifrangere le onde provenienti da un oggetto. La rifrazione è il meccanismo in base al quale vengono messi a fuoco gli stimoli luminosi. Il cristallino, rispetto alla cornea, ha maggiore potere. Si dice che ha maggior capacità di accomodazione, proprio per sottolineare che grazie a questo sistema di muscolatura presente nel cristallino, questo ha maggior potere di modifica strutturale che permette la messa a fuoco degli oggetti. I raggi che passano lontani dal centro del cristallino determinano un'immagine meno nitida e quindi ridurre il diametro della pupilla serve a migliorare la nitidezza della percezione. I difetti al sistema diottrico (come la miopia) sono facilmente e parzialmente correggibili, mentre problemi alla retina sono molto più seri e rilevanti. Abbiamo detto che il disco ottico è la porzione essenziale perchè sia di ingresso che di uscita dalla retina. È una porzione che non ha recettori, unica zona della retina con questa caratteristica. Il disco ottico è responsabile del fenomeno del punto cieco (macula cieca). Abbiamo due regioni, corrispondenti alle porzioni nasali, zone essenzialmente monoculari, mentre l'altra porzione, temporale, riceve le informazioni da entrambi gli occhi. Il fenomeno della macula cieca si può associare al fatto che con la chiusura di un occhio e la fissazione di uno dei due spot in modo continuativo, si vedrà come non si riesce a percepire l'altro spot che inizialmente si vedeva. Questo succede perchè si tratta di una proiezione che chiama in causa la porzione nasale dell'occhio aperto ed essenzialmente si scontra con la presenza di una zona d'ombra, priva di fotorecettori, che non permette di vedere stimoli al di fuori della porzione di campo visivo che viene esaminata dall'occhio che resta aperto. Un'altra caratteristica è che nella fovea sono presenti soltanto i coni. I fotorecettori reticini si divono in due categorie diverse: coni e bastoncelli. I coni sono presenti nella fovea, oltre che nel resto della retina. Nella fovea ci sono solo coni e questo avrà una sua implicazione. La retina nasce dalla vescicola ottica, una sorta di invaginazione del telencefalo. Questa vescicola, nei passaggi successivi dello sviluppo, forma il calice ottico e questo poi avrà due porzione, una interna, quella che origina la retina vera e propria, una esterna che origina l'epitelio pigmentato. Questo contiene melanina ed è una sostanza importante per ridurre la dispersione della luce che penetra nell'occhio. Subito al di sotto dell'epitelio pigmentato abbiamo le cellule recettrici, dette fotorecettori, recettori di III ordine, perchè non sono direttamente connessi a neuroni sensitivi primari, ma tra il recettore e il neurone sensitivo primario c'è una serie di interneuroni. I corpi cellulari di questi recettori si trovano al livello dello strato nucleare esterno. I fotorecettori fanno contatto con degli interneuroni. I punti di contatto sono a livello dello strato plessiforme esterno, mentre i corpi cellulari dei neuroni sono nello strato nucleare interno. Questi interneuroni hanno tre nomi diversi perchè hanno funzione e morfologia diversa: cellule bipolari, cellule amacrine, cellule orizzontali. Gli interneuroni contattano a loro volta a livello dello strato plessiforme interno le cellule ganglionari, che hanno i corpi cellulari situati nello strato delle cellule gangliali e i neuroni sensitivi primari inviano contatti che escono dalla retina tramite il disco ottico e le fibre proseguono per il disco ottico fino al sistema nervoso centrale. L'epitelio pigmentato è importante anche perchè mantiene l'intero apparato di fototrasduzione, perchè ha due funzioni principali: la fagocitosi dei dischi esausti. Sia i dischi che i fotopigmenti sono elementi che costituiscono una prozione del recettore, sia del cono che del bastoncello. L'epitelio pigmentato è connesso a una porzione più esterna dei fotorecettori. Questa è costituita da una sorta di dischi membranosi, che vengono continuamente riciclati, quindi c'è un pezzo di recettori che viene continuamente riciclato e smaltito in questo modo: si increspano, poi si ha un'estremità che si separa dal recettore. Quest'estremità che dev'essere riclicata, viene inglobata dall'epitelio pigmentato che sta sopra.Vediamo come sono costituiti i fotorecettori. La morfologia è diversa da coni e bastoncelli. I bastoncelli sono più allungati. Gli elementi essenziali però sono gli stessi: segmento esterno, che nel caso del bastoncello è costituito da dischi di membrana e nel caso del cono sono più introflessioni ed estroflessioni di membrana. Entrambe queste parti sono dette segmenti esterni. Questi sono porzioni in cui la luce viene trasformata in segnale elettrico; è la componente essenziale per cominciare il processo di fototrasduzione. Le altre porzioni sono il segmento interno e la terminazione sinaptica. Il segmento interno comprende il nucleo, i mitocondri e le zone principali delle cellule per la sintesi proteica. Viceversa, la terza porzione, la terminazione sinaptica, cioè l'assone del fotorecettore, che fa contatti sinaptici con i dentriti dei neuroni. La categoria di interneurone viene comunemente detta categorie delle cellule bipolari, anche se di fatto comprendono i tre sottotipi di cellule bipolari, amacrine e orizzontali. Il processo di fototrasduzione inizia con un fotone che viene assorbito da parte del fotopigmento contenuto nei dischi di membrana o nelle estroflessioni di membrana. Il fotopigmento è quindi contenuto nel segmento esterno del recettore. La porzione chiamata pigmento visivo è costituita da un fotopigmento che contiene una proteina detta retinale, che è un derivato della vitamina A, e una molecola della famiglia delle opsine. Il retinale è la porzione di fotopigmento che assorbe la radiazione, ma dev'essere sempre associato all'opsina per avere una modifica strutturale legata all'ingresso della radiazione. L'insieme retinale-opsina viene modificato strutturalmente una volta che arriva un fotone e questa modifica che subisce è responsabile della cascata successiva a livello molecolare. Questa prevede che si abbia un'attivazione di un secondo messaggero chiamato proteina G trasducina. A sua volta, questa trasducina è responsabile dell'attivazione di una fosfodiesterasi e questa è responsabile dell'idrolisi del GMP ciclico. Il passaggio fondamentale sta proprio nel riuscire a smaltire (idrolizzare) le molecole di GMPc presenti a questo livello legate alle possibilità che si hanno di ingresso nel fotorecettore attraverso il canale presente a questo livello. Al buio c'è un canale sensibile agli ioni sodio che permette l'ingresso di GMPc che è presente in grande quantità all'interno del segmento esterno del fotorecettore. Si ha una depolarizzazione del potenziale a membrana, cioè in condizione di riposo. Quando arriva la luce, questa colpisce il pigmento visivo, o meglio il complesso retinale-opsina, che cambia conformazione. Questo cambio di conformazione provoca l'attivazione di tutta la cascata intracellulare, quindi si ha attivazione del messaggero intracellulare (trasducina). Questa a sua volta attiva la fosfodiesterasi che, a sua volta, causa l'idrolisi del GMPc e, come ultima conseguenza, il canale per il sodio sensibile alla presenza di GMPc, in assenza di esso, si chiude e gli ioni sodio non entrano più. Si passa a una iperpolarizzazione del potenziale di membrana. L'arrivo di uno stimolo visivo ha quindi come effetto finale l'iperpolarizzazione, cosa anomala, perchè nella maggior parte degli altri sistemi uno stimolo causa una depolarizzazione. Un'altra caratteristica fondamentale di questo processo è il fatto che c'è un'amplificazione molto forte dell'effetto. Da un solo fotone che arriva al pigmento visivo, si ha una risposta che arriva a coinvolgere l'attività di almeno centomila molecole di GMPc libere nello spazio del segmento esterno. Questo è la prima modalità con cui lo stimolo visivo arriva al recettore. L'altra caratteristica interessante è che la luce, di per sè, serve solo a operare il primo tipo di modifica strutturale. Tutto ciò che avviene dopo è opera della cascata, ma non è più mediata dallo stimolo visivo. L'opsina del bastoncello è solo e sempre di un tipo: rodopsina. Viceversa, i coni hanno tre tipi di opsine diverse. Tra le altre differenze funzionali: i bastoncelli sono essenziali in condizioni di scarsa illuminazione (visione notturna o scotopica). Essi hanno quantità di pigmento, permettono maggiore amplificazione del segnale e hanno connessioni convergenti. Di base, le connessioni convergenti hanno come conseguenza il fatto che diminuisce l'acuità di un sistema. Viceversa, i coni che hanno connessioni non convergenti, hanno un'altissima acuità visiva e quindi altissima capacità di discriminare i dettagli. I bastoncelli non sono presenti nella fovea, ma soltanto i coni. Viceversa, i coni non sono presenti nelle porzioni più periferiche della retina. Nella retina abbiamo circa 90 milioni di bastoncelli e 4 milioni e mezzo di coni. La densità di coni e bastoncelli è diversa nelle diverse porzioni della retina. I coni sono rappresentati nelle porzioni più centrali e i bastoncelli nelle porzioni più periferiche. Un esempio di convergenza è il fatto che più bastoncelli convergono sugli stessi interneuroni. Il tratto principale della convergenza è il fatto che abbiamo più bastoncelli sullo stesso interneurone. I coni hanno connessioni più privilegiate. Oltre ad avere diversi tipi di connessioni, si distinguono due tipi di sistemi: orizzontali e verticali. Il sistema orizzontale è dato da un recettore che ha connessioni con i diversi tipi di interneuroni e con le cellule ganglionari e nel sistema verticale il recettore ha connessione eslusiva con una cellula bipolare e con la cellula ganglionale. Questi due sistemi danno indicazione sui diversi tipi di complessità di processamento sulla retina. Quello orizzontale ha infatti una funzione aggiuntiva che è quella di portare con sè le informazioni degli altri interneuroni. Un'altra caratteristica essenziale è che i fotorecettori e gli interneuroni non generano potenziale d'azione. Il processo di iperpolarizzazione non porta a un potenziale d'azione ma a un potenziale di recettore, trasmesso agli interneuroni, ma non si ha generazione di potenziale d'azione, ma di potenziali elettrotonici. Solo le cellule ganglionari generano potenziale d'azione. L'intero processo attiva potenziali di recettori e solo se sarà soprasoglia sarà in grado di attivare la cellula membrionale e di far innescare un potenziale d'azione in questa cellula. Questo avviene anche per una ragione di spazio. Questo processo avviene in una porzione limitata, nella retina. Non c'è necessità di inviare l'informazione a lunghe distanze, come nel sistema tattile, in cui è necessaria un'amplificazione notevole del segnale. Abbiamo la suddivisione delle cellule ganglionari in due tipologie: cellule magnocellulari e cellule parvocellulari. Le prime sono quelle che hanno campi recettivi più grandi e sono responsabili dell'analisi del movimento; le parvocellulari hanno campi recettivi più piccoli e ricevono informazioni su forma e colore. Il sistema M fa capo alle cellule magnocellulari e il sistema P alle parvocellulari, che hanno funzioni diverse nell'analisi dello stimolo visivo. La scomposizione e analisi dello stimolo visivo avviene quindi già a livello della retina. Ci soffermiamo sulle cellule bipolari. L'interneurone ha un campo recettivo di grandezze diverse a seconda della porzione di retina che rappresenta, caratterizzato da zone eccitatorie e zone inibitorie disposte in modo circolare. I campi recettivi che sono, in questo caso, l'area della retina la cui stimolazione luminosa comporta una modifica dell'attività elettrica della cellula, hanno dimensioni diverse (nella fovea i campi sono piccolissimi, così come sulla punta delle dita si ha massima acuità tattile e minima dimensione e massima densità dei recettori). Per attivare queste cellule lo stimolo migliore sarà uno stimolo che darà un'informazione maggiore per il contrasto. A differenza dei fotorecettori che hanno stimoli che corrispondono a differenti intensità luminose, qui l'intensità non è più importante, lo è l'intensità luminosa e il contrasto. Tutto questo vale per le cellule bipolari e ganglionari. Lo stimolo che arriva al centro attiverà anche due diversi tipi cellulari successivi: le bipolari che hanno centro on e le ganglionari che hanno centro on. L'iperpolarizzazione del recettore di fatto comporta una diminuzione di rilascio di neurotrasmettitore nello spazio sinaptico. Le cellule gangliari non sono mai silenti, neanche al buio, ma la loro frequenza di scarica è modulata dallo stimolo luminoso. La loro frequenza di scarica non è proporzionale all'intensità assoluta dello stimolo, ma piuttosto alla differenza di intensità esistente fra il centro e la periferia del campo recettivo. Quelle a centro ON rispondono meglio quando è illuminato il centro e non la periferia, l'opposto quelle a centro OFF, analizzando di fatto i contrasti piuttosto che la luminosità assoluta. La luminosità è influenzata dallo sfondo perché le nostre cellule retiniche riescono a visualizzare i contrasti tra centro e periferia, oggetto e sfondo. Le cellule ganglionari segnalano bene anche le variazioni di intensità luminosa: - se l’intensità luminosa aumenta nel centro del campo recettivo di una cellula ganglionare a centro ON, la sua frequenza di scarica di potenziali d’azione aumenta; - se l ’intensità luminosa diminuisce nel centro del campo recettivo di una cellula ganglionare a centro OFF, la sua frequenza di scarica di potenziali d’azione aumenta. Questo principio di analisi di base con cui riusciamo a percepire i contrasti di intensità luminose è lo stesso con cui organizziamo anche la nostra esplorazione delle scene visive. Sono stati condotti tanti studi di registrazione dei movimenti oculari e in particolare quando si elaborano stimoli complessi come i volti si è visto come l'esplorazione oculare si concentra su quelle zone particolarmente importanti per i contrasti, come bocca, naso e occhi. 14. Sistema visivo 2 Suddiviamo il campo visivo in due tipi di quadranti. Immaginiamo di tirare una linea che divida la porzione destra e la porzione sinistra del campo visivo. Si possono suddividere ulteriori quadranti, ma essenzialmente ci riferiamo a stimoli che cadono nella porzione destra o sinistra del campo visivo. Uno stimolo nella porzione destra del campo visivo viene proiettato e codificato dall'emiretina nasale dell'occhio dell'occhio di destra e dall'emiretina temporale dell'occhio di sinistra. La retina ha quindi queste due suddivisioni: emiretina nasale (più vicina al naso) ed emiretina temporale (più vicina ai lati). Viceversa per gli stimoli del campo visivo sinistro. La rappresentazione è capovolta, quindi bisogna pensare a una suddivisione in altezza dello stimolo. Se esso cade nella parte superiore del campo visivo viene proiettata nelle porzioni inferiori della retina e viceversa. Di base, ogni stimolo arriva a entrambi gli occhi anche se in porzioni diverse della retina. Il fatto che arrivi a entrambi gli occhi è anche la ragione per cui non ci accorgiamo normalmente del punto cieco (si è detto che il punto cieco è quella zona in cui non ci sono fotorecettori). Le fibre che escono tramite il disco ottico dalle retine incrociano – ma non tutte, una parte – nel chiasma ottico. Si incrociano soltanto quelle fibre che corrispondono alle emiretine nasali, andando a convergere nel tratto ottico controlaterale. Il chiasma ottico è la zona essenziale per la nostra visione binoculare. Nel caso di animali che hanno visioni laterali come il cavallo le fibre provenienti da un singolo occhio incrociano tutte le fibre nel chiasma ottico. Il tratto ottico di sinistra alla fine quindi conterrà le fibre provenienti dal settore temporale della retina dell'occhio sinistro e le fibre provenienti dal settore nasale della retina dell'occhio destro. Sono comunque fibre che si riferiscono alla stessa porzione di campo visivo. Di base, a ciascun tratto ottico arrivano informazioni che fanno riferimento alla stessa zona di campo visivo. Quasi la totalità del campo visivo ha una visione binoculare, ma ci sono porzioni più estreme e laterali del campo visivo di destra e sinistra che hanno una proiezione monoculare, cioè arriva in modo privilegiato alla porzione nasale dell'occhio ipsilaterale. Generalmente quest'informazione monoculare in assenza dell'attività dell'altro occhio ci fa sperimentare il fenomeno di punto cieco. Le fibre che escono da ciascun chiasma ottico sono detti nervi ottici. La successiva stazione è talamica (quindi siamo a livello sottocorticale) ed è chiamata corpo genicolato laterale. A questo livello arrivano fibre che permettono di costruire la stessa porzione di campo visivo ed è una ricostruzione che viene fatta in modo controlaterale. Se avessimo una lesione a livello del nervo ottico, avremo un caso di cecità, proprio perchè sono lese le fibre iniziali. In letteratura sono descritti molti casi di lesioni al chiasma ottico, che hanno conseguenze diverse in base alle porzioni specifiche che vengono colpite nel chiasma ottico. Generalmente i danni dovuti a lesione di chiasma ottico vengono chiamati emianopsia perchè hanno sempre come conseguenza primaria una perdita della visione di metà del campo visivo. Queste vengono ulteriormente specificate con i termini di temporale o nasale a seconda se riguardano fibre della porzione temporale o nasale della retina. Parliamo di emianopsia omonime o eteronome a seconda che riguardino la stessa porzione di campo visivo o porzioni diverse, in particolare quelle omonime sono legate a regioni di tratti ottici perchè abbiamo visto che nel tratto ottico le fibre che arrivano sono quelle che già danno informazione dalla stessa porzione di campo visivo. Ci possono essere emianopsie anche in seguito a lesioni della stazione talamica e stazioni sottocorticali successive. Ma se la lesione coinvolge entrambi gli emisferi si ritorna a parlare di cecità. Dal talamo, attraverso la radiazione ottica, si arriva alla corteccia visiva. Gli assoni terminamo nel quarto strato della corteccia. La corteccia visiva primaria è classicamente definita come V1 o come area 17 di Brockman. Si trova essenzialmente nel polo occipitale del nostro emisfero. Tuttavia, una mole indefinita di studi ha dimostrato che le aree visive sono molto più estese della corteccia visiva primaria; si contano addirittura 36 aree. Attualmente è noto che l'elaborazione dello stimolo visivo coinvolge l'intera corteccia visiva nel polo occipitale, ma anche varie porzioni del polo temporale e parietale di entrambi gli emisferi. È detta corteccia striata la V1 perchè gli assoni che terminano nel quarto strato formano la stria di Gennarini, con morfologia striata. Tutta la via viene detta retino-stiata proprio perchè dalla retina le informazioni arrivano ai neuroni corticali della corteccia visiva primaria. Questa non è l'unica esistente per l'analisi degli stimoli visivi. Le altre vie partono dai tratti ottici. Il 90% delle fibre di un tratto ottico converge nella via retino-striata, ma ci sono proiezioni anche ad altre tre strutture: il pretetto (regione nel mesencefalo), il collicolo superiore e il nucleo soprachiasmatico dell'ipotalamo. L'ultima serve per regolare i ritmi circadiani, quella pretettale serve per controllare il riflesso di aumento o diminuzione del diametro della pupilla rispetto alle condizioni di illuminazione e la via mediata dal collicolo superiore ha una funzione di orientamento verso uno stimolo visivo (serve cioè per dirigere il movimento oculare verso un oggetto in risposta a stimoli visivi). La via pretettale è responsabile del riflesso pupillare e si dice che è un riflesso consensuale, che coinvolge cioè i due occhi nella stessa misura. Se il riflesso non fosse consensuale significherebbe che c'è una patologia di questo sistema. È un'area del mesencefalo collegata sia al sistema parasimpatico che al sistema ortosimpatico; questi collegamenti formano una sorta di circuito perchè dalla retina, tramite il tratto ottico, l'informazione arriva all'area pretettale, si ha l'attivazione del sistema parasimpatico e tramite il terzo nervo cranico vengono stimolate altre cellule situate a livello del ganglio cigliare. Queste coordinano il movimento di contrazione e dilatazione della muscolatura dell'iride; in particolare, in caso di attivazione del parasimpatico, si ha attivazione dei muscoli che restringono l'iride causando diminuzione del diametro della pupilla in condizioni di luce molto intensa (miosi). Il riflesso opposto, che si ha in condizioni di luce debole, è legato al fatto che da quest'area pretettale le informazioni arrivano all'ortosimpatico e a loro volta riconvergono sulla muscolatura dell'iride causando dilatazione del diametro pupillare (midriasi). Le cellule ganglionari a capo di questa via sono particolari, perchè sono una sottocategoria di cellule che riescono a rispondere, quindi aumentare la loro frequenza di scarica, in risposta a variazioni di intensità luminosa e non in risposta a contrasto di luminosità (a cui rispondono il 90% di cellule ganglionari). Il collicolo è situato a livello del mesencefalo. È una stazione importante perchè qui convergono diverse modalità sensoriali. È formata da strati diversi, in totale 7, ma di base all'interno di questi strati sono contenute tre mappe sensoriali principali: mappa per la visione, situata negli strati superiore, mappa per il tatto e mappa per l'udito nella porzione inferiore del collicolo. Tutte queste mappe sono connesse con quella motoria e questo spiega come facciamo a organizzare le reazioni agli stimoli. Anche il nucleo genicolato del talamo laterale è suddiviso in 6 diversi strati. Ci sono due caratteristiche importanti: le afferenze di entrambi gli occhi sono segregate, cioè ci sono strati che ricevono solo da un occhio e altri che ricevono dall'altro, ma mai all'interno di uno stesso strato convergono afferenze di entrambi gli occhi, anche se di fatto tutto il corpo genicolato laterale riceve l'informazione della stessa porzione di campo visivo. L'altra suddivisione è quella in strati M e P. Ieri si sono viste le cellule magno e parvocellulari. Gli stati M (primi due della stazione talamica) ricevono le afferenze dalle gangliari di tipo M. Le cellule di tipo M codificano per il movimento. Gli altri strati P sono quelli che vanno dal 3 al 6 e ricevono dalle cellule gangliari di tipo P, deputate all'analisi della forma e del colore. Questa precisa e dettagliata organizzazione a livello della stazione talamica fa sì che già a questo livello sottocorticale ci siano delle mappe, rappresentazioni retinotopiche. Esse sono disposte a registro, cioè ciascun strato presenta una mappa con dimensioni sproporzionate com'è caratteristica generale delle mappe sensoriali. In questo caso le porzioni più rappresentate sono quelle che ricevono le informazioni dalle zone foveali, ad esempio quelle hanno maggiore densità e da campi recettivi più piccoli avranno dedicata una porzione di mappa maggiore. Al quarto strato arrivano sempre informazioni da un solo occhio, quindi l'informazione arriva segregata, così come nel talamo. Gli altri strati, sovrastanti o sottostanti al IV strato, codificano informazioni da entrambi gli occhi, anche se all'interno di ciascuna regione topografica della corteccia visiva ci sono neuroni che risponderanno in modo privilegiato a un occhio rispetto ad un altro. La mappa è presente sia a livello del talamo che a livello della corteccia visiva primaria. Parliamo di mappa retinotopica, che rappresenta le diverse regioni della retina che codificano per diverse zone del campo recettivo. Le porzioni codificate dalle regioni foveali sono quelle che occupano maggiori spazi sulle due cortecce visive. La corteccia visiva primaria è situata nella porzione sovra e sottostante della scissura calcarica, solco che separa le due porzioni di corteccia. È una mappa sproporzionata, ma proporzionale alla densità dei recettori e di cellule ganglionari presenti nelle diverse regioni della retina. La fovea è rappresentata su 50% della corteccia, proprio perchè la nostra visione dipende in gran parte dall'informazione codificata dalla fovea. Il corpo genilato laterale contiene anche informazioni che vanno dalla corteccia ai sistemi discententi, al contrario. Infatti gran parte delle fibre presenti a livello della stazione talamica del corpo genicolato laterale proviene in realtà dalla corteccia e soltanto una parte minima sono le fibre che da questa stazione alla corteccia. La funzione di questa fibre è quindi quella di inbire il processo corticale di una serie di stimoli visivi presenti nel nostro campo ma che non elaboriamo a livello di consapevolezza cosciente. Vediamo ora quali sono le cellule della corteccia e che campi recettivi hanno. I campi recettivi sono costituiti da zone circolari con zone eccitatorie e inibitorie disposte al centro e in periferie. Questo è stato precedentemente visto per le cellule ganglionari e neuronali ed è stato dimostrato essere così anche per le cellule talamiche, della stazione sottocorticale. Il problema è che quando per la prima volta gli scienziati hanno iniziato a registrare con i microelettrodi l'attività dei neuroni della corteccia, ad esempio degli strati sovra e sottostanti al IV strato, hanno visto che mettendo la scimmia di fronte allo schermo nero col puntino bianco che si spostava in varie direzioni, nessuno di questi neuroni rispondeva e di fatto si comportavano in modo diverso rispetto alle altre cellule di elaborazione dello stimolo. Un giorno succede che si va ad incastrare la diapositiva che doveva mandare lo spot e l'immagine che si proietta sullo schermo non è quella di un puntino ma di una linea. Le cellule iniziano a rispondere. La scoperta risulta che lo stimolo migliore a livello corticale non è un punto, ma dev'essere allungato, con un orientamento particolare, come una linea, e che abbia una posizione precisa all'interno dello spazio. A questo punto sono state studiate le diverse categorie cellulari presenti in corteccia. Le categorie principali sono due: cellule semplici e cellule complesse. Le cellule semplici hanno campo recettivo dato quindi dalla somma di campi recettivi centro on e centro off, ma è un campo recettivo attivato da stimoli che hanno aspetto allungato e orientamento caratteristico. Quando la barra viene posta orizzontalmente, le cellule non rispondono, mentre a seconda che la barra vada a colpire più o meno, in base al suo orientamento, le porzioni eccitatorie del campo recettivo, allora ci sarà aumento della frequenza di scarica, con apice massimo nella circostanza in cui la barra di luce va a cadere esattamente nella porzione eccitatoria del campo recettivo della cellula. L'informazione sul contrasto di luminosità nel campo recettivo, generata dalle cellule ganglionari e talamiche, è mantenuta dalle cellule semplici ed incorporata in un'informazione più complessa sulla forma e sull'orientamento dell'oggetto. Da questo si vede che aumentato la gerarchia corticale, aumenta anche la complessità. Nelle cellule complesse non ci sono più zone inibitorie ed eccitatorie, centri on e off. Si pensa che la costruzione del campo recettivo sia data dalla somma di campi recettivi di cellule semplici. Gli stimoli adeguati per queste cellule sono di nuovo linee dotate di orientamenti particolari. La linea posta orizzontalmente non attiva queste cellule. Le linee che cambiano gradualmente orientamento e poi vengono disposte parallelamente aumentano gradualmente la loro frequenza di scarica, ma anche quando lo stimolo è posto nelle porzioni più laterali, c'è la massima risposta della cellula. Queste dunque segnalano il concetto astratto di orientamento, senza più un particolare riferimento alla posizione. Alcune rispondono meglio a barre luminose orientate che si muovono attraverso il campo recettivo. Sembra persa, da un lato, l'informazione relativa alla posizione spaziale dello stimolo, ma di fatto questo è un esempio di analisi mirata a localizzare l'oggetto in senso lato, non in modo preciso. Quello che conta è perciò il riconoscimento dello stimolo e non tanto più la sua esatta posizione nello spazio. A mano a mano che si procede nella gerarchia corticale, ci sono stazioni che sono responsabili di informazioni con caratteristiche sempre diverse e complesse per poter riconoscere e identificare lo stimolo. Tutte queste cellule danno informazioni sui contorni dell'oggetto, permettendo di ricostruire le sue forme. Due ulteriori concetti importanti nel sistema visivo sono quelli di colonne di dominanza oculare e di colonne di orientamento. Tutti gli studi sulla corteccia cerebrale hanno contribuito a comprendere, da un lato, qual è l'attività funzionale delle cellule che la compongono e, dall'altro, com'è organizzata strutturalmente. Si è scoperto che ci sono settori di corteccia chiamate colonne, dominate in modo specifico dalle afferenze provenienti da un occhio. I neuroni sono segregati nel senso che ricevano informazioni segregate da un occhio e non dall'altro, come abbiamo detto, a livello del IV strato, mentre negli altri no. Ci sono colonne però in cui prevale l'informazione proiettata da un occhio. La scoperta di queste colonne è stata fatta registrando i neuroni presenti in ciascuna colonna e si è visto come si tratta di neuroni che rispondono preferenzialmente a un occhio rispetto a un altro. Nelle colonne di orientamento i neuroni rispondono in modo privilegiato a orientamenti specifici degli stimoli. Vi è un'interazione ortogonale a livello della corteccia tra colonne di dominanza e colonne di orientamento, tant'è che se immaginiamo di guardare un pezzo di corteccia possiamo vedere che ci sono dei blocchi identificabili chiamate ipercolonne. Ogni ipercolonna rappresenta un pezzo di corteccia che codifica per una ragione di retina precisa e avrà un orientamento e dominanza oculare per un determinato occhio. L'ipercolonna deriva dall'insieme di questi due sistemi e quindi ci saranno colonne che risponderanno in modo privilegiato a un occhio rispetto che a un altro e a stimoli con orientamento preciso rispetto che ad altri. Mescolati a questo sistema di colonne ci sono altre colonne dette blob. Queste forme inserite all'interno sono delle ulteriori colonne responsabili essenziali dell'analisi del colore. Si dice che ogni ipercolonna avrà delle zone blob e delle zone interblob. Le prime sono quelle in cui sono inseriti i blob e le seconde sono quelle in cui non sono presenti blob, quindi sono porzioni di ipercolonna responsabili dell'analisi della forma e del movimento. Ricapitolando: le cellule ganglionari, retiniche e talamiche rispondono a contrasti di luminosità, maggiori ai bordi. Le cellule della corteccia V1 elaborano quest'informazione sul contrasto, ma aggiungendo l'orientamento e i bordi sono ancora l'informazione più significativa attraverso il quale andiamo a costruire le forme dell'oggetto. I neuroni a livello corticale sono specializzati nel riconoscimento di forme specializzate come quelle dei volti, tramite un'analisi che avviene in modo gerarchico dagli stadi retinici fino agli stadi corticali. 15. Sistema visivo 3 La percezione dei colori è una caratteristica del nostro sistema visivo. Ci sono animali che non vedono i colori, ma solo in bianco e nero. Pensando a sistemi visivi che non sono in grado di percepire il colore, si può dire che le differenze fra un oggetto e l’altro vengono codificate come differenze di intensità luminosa e quindi differenze di quantità di luce che viene riflessa da un oggetto e non da un altro, parlando sempre in termini assoluti di intensità. Con il colore subentra un altro elemento che è quello delle lunghezze d’onda. Percepiamo due oggetti di colore diverso perché ciascuno di essi riflette determinate lunghezze d’onda e non altre. Riuscire a vedere a colori, quindi riuscire a discriminare quali sono le lunghezze d’onda riflesse da un oggetto e non da un altro favorisce la nostra percezione di contrasto tra gli oggetti. L’informazione sul contrasto è una delle più salienti per quanto riguarda il sistema visivo proprio perché abbiamo anatomicamente, strutturalmente delle cellule che a diversi livelli, dalla retina fino alla corteccia rispondono ai contrasti. Quindi, un’informazione aggiuntiva che aumenta l’importanza del contrasto come quella del colore porta all’apice massimo le potenzialità del nostro sistema visivo. In che modo noi riusciamo a vedere a colori? Anatomicamente, strutturalmente quali sono i correlati di questa visione cromatica? Uno dei primi esperimenti, che ha voluto indagare quali sono i meccanismi nella nostra percezione dei colori, è stato un esperimento fatto da Newton. L’esperimento classico è stato quello di proiettare un fascio di luce bianca attraverso un prisma di vetro e sperimentalmente si è osservato che la luce bianca all’uscita dal prisma non è più bianca ma dà tutta la gamma dei colori proprio perché si rifrange sul prisma e all’uscita le onde di diversa lunghezza d’onda appaiono di colore diverso. Questo primo esperimento faceva pensare che la percezione del colore fosse proprio legata alla lunghezza d’onda che l’oggetto è in grado di riflettere ogni volta. Tuttavia, pensando alla nostra esperienza comune non è assolutamente così, perché altrimenti ogni volta che cambia la fonte di illuminazione, che è una delle caratteristiche che fa variare anche la lunghezza d’onda riflessa da un oggetto, allora il colore dell’oggetto dovrebbe variare. Viceversa, sappiamo che ogni volta che cambiano le condizioni di illuminazione il colore resta sempre costante. Il colore di un limone è sempre giallo sia sotto il sole sia sotto la luce elettrica, quindi non è soltanto questo che può spiegare come mai riusciamo a vedere i colori. Un altro esempio facile da realizzare è mettere lo stesso oggetto, ad esempio, un cubo con tanti pezzettini colorati, sotto sorgenti luminose che hanno filtri diversi per la luce gialla, bianca o blu. Considerando il colore dei quadratini che compongono la faccia superiore del cubo, noi li vediamo di colore costante nonostante cambi la sorgente di illuminazione. I quadratini rossi sotto la luce gialla continuano ad essere visti come quadratini rossi e così anche sotto la luce bianca o sotto la luce blu. Di fatto il nostro sistema visivo si fonda su una proprietà importantissima e cioè la proprietà della costanza dei colori. Questa proprietà spiega anche biologicamente il significato stesso della visione cromatica perché, se non avessimo questo principio organizzatore e quindi la nostra percezione dei colori variasse di volta in volta in base alla sorgente luminosa, non avrebbe neanche più senso vedere a colori, perché il colore sarebbe una cosa del tutto opinabile, che cambia ogni volta che cambia la sorgente. Viceversa per il nostro sistema è importantissimo vedere a colori perché aumentiamo i contrasti e quindi aumentiamo al massimo le potenzialità di elaborazione dei nostri neuroni, ma il colore deve essere un tratto costante. La costanza dei colori a che cosa è legata? Il motivo della costanza cromatica è legato al fatto che facciamo dei confronti tra oggetti vicini, o tra oggetto e superficie, oggetto e sfondo. Il confronto è un confronto di riflettanze. Le riflettanze sono le quantità di onde di diverso colore che un oggetto riesce ad emettere. La riflettanza è proprio un’espressione quantitativa dell’emissione delle lunghezze d’onda di ogni oggetto. Ogni oggetto ha una riflettanza che lo caratterizza, cioè riesce a riflettere un quantitativo fisso di onde elettromagnetiche, ed è il confronto tra le riflettanze di due oggetti diversi o di un oggetto con la sua superficie sfondo che ci permette di mantenere costanti i colori. Due esperimenti: il colore di un oggetto non viene giudicato solo analizzando le lunghezze d’onda che riflette, ma paragonando le quantità di raggi di lunghezze d’onda diverse riflesse dall'oggetto con quelle riflesse dallo sfondo (o da oggetti circostanti). Il colore diventa in questo modo una caratteristica qualificante l’oggetto ma anche il colore è una percezione che abbiamo da un meccanismo di confronto che opera il nostro sistema nervoso. Questa teoria è detta Teoria “retinex” di Land perché Land è stato il pioniere di queste ricerche (oltre ad essere il padre della Polaroid). L’esperimento chiave di Land viene chiamato esperimento di Land-Mondrian (fine anni ‘70) perché la configurazione stimolo che ha usato Land è simile a un quadro di Mondrian. Nell’esperimento ci sono due quadri Mondrian o comunque due stimoli di tipo Mondrian posti uno accanto all'altro e ci sono tre proiettori diversi. Questi tre proiettori hanno dei filtri, uno per la luce rossa, uno per la luce verde e uno per la luce blu. Il tipo di luce che proiettano sullo stimolo, che è il Mondrian, è diverso come colore iniziale. Questi tre proiettori sono dotati di reostati che riescono a far variare anche l'intensità della luce proiettata sul Mondrian. Tre luci regolabili: quando si parla di lunghezza d'onda corta si parla di luce che appartiene allo spettro del blu (quando si guarda tutto lo spettro delle lunghezze d’onda della luce visibile, quelle che hanno lunghezze d'onda più corte sono quelle che noi percepiamo come blu), le lunghezze d’onda intermedie sono quelle che percepiamo come verdi e quelle dalle lunghezze d'onda più lunghe sono quelle che percepiamo come rosso o rosso profondo. Tre luci regolabili, lunghezze d'onda corta, media e lunga, e vanno ad illuminare questa serie di cartoncini colorati. Ad esempio l'autore si era soffermato su un cartoncino verde vicino a quello marrone ed era andato ad illuminare il cartoncino verde in modo tale che andasse poi a riflettere la stessa lunghezza d'onda di quello marrone, ma ha visto che la sua percezione era comunque di un colore verde costante e di un colore marrone costante. Di fatto, anche se interveniamo sperimentalmente sulla fonte e sul tipo di onda che riesce poi ad emettere l'oggetto, il colore non cambia. Quindi, grazie agli esperimenti di Mondrian si comprende che la costanza del colore non è legata né alla sorgente di per sé, né alla quantità di lunghezze d'onda che vengono emesse dall'oggetto ma dipende proprio dalla costanza delle riflettanze di oggetti diversi. Questo concetto l’ha tirato fuori con due altri esperimenti in cui ci sono sempre due configurazioni Mondrian. Nel primo esperimento Land ha usato tre proiettori con un'intensità luminosa di 100 mV per ciascun proiettore. Nel grafico vediamo l'intensità di ciascuna delle tre sorgenti, la percentuale di riflettanza (la quantità di lunghezze d'onda che ciascuno dei due Mondrian riesce a riflettere per quei tre determinati colori, per quelle tre fonti specifiche di intensità luminosa). Land ha registrato quello che succede nel primo e nel secondo Mondrian con queste condizioni iniziali di illuminazione. Si tratta di configurazioni leggermente diverse quindi i colori all'interno di ciascun Mondrian e gli oggetti erano mescolati in modo diverso. Poi ha modificato l'intensità delle luci provenienti dai proiettori quindi 300 mW per il rosso, 400 per il verde, 140 peril blu e ha registrato di nuovo le riflettanze delle due superfici tra loro. Quello che ha notato è che il primo Mondrian emette con intensità 90 nel rosso e il secondo Mondrian sempre nel rosso con intensità 30: quindi ha fatto la seguente proporzione 90 sta a 30 come 270 sta 90 nel caso della seconda configurazione. Di fatto ottiene sempre lo stesso tipo di rapporto tra le riflettanze delle superfici in questo caso e le riflettanze delle superfici in quest'altro caso. In questo modo Land è andato a vedere che anche se aumento l'intensità della sorgente su due configurazioni stimolo che sono tra loro diverse come colori, di base percepirò sempre costanti i colori di queste configurazioni perché i rapporti fra le riflettanze di queste due superfici restano costanti. Quindi può cambiare la lunghezza d'onda che emette ogni superficie sulla base dell’intensità di sorgente luminosa ma non cambierà mai il rapporto tra la sua riflettanza e quella della superficie accanto. Ed è questo rapporto, questo confronto tra superfici diverse, oggetti vicini, tra una sola superficie e il suo sfondo che ci permette di percepire i colori costanti. Se facciamo invece riferimento alle due facce di un cubo, in realtà possiamo vedere come non abbiamo, in questo caso, costanza di colore se consideriamo pezzettini del cubo che appartengono alle due facce, perché in questo caso specifico il nostro confronto viene operato tra i quadratini di ogni superficie, quindi noi riusciremo a percepire sempre quelli rossi come rossi, quelli verdi come verdi, quelli bianchi come bianchi, perché stiamo operando un confronto di tutti gli oggetti, quindi delle riflettanze di tutti gli oggetti posti su questa cosa configurazione della faccia superiore. Quando invece passiamo ad esaminare quest’altra faccia del cubo noi andiamo a discriminare il colore di questi quadratini sulla base del confronto delle riflettanze di questi quadratini. In questo caso è come se avessimo due stimoli indipendenti che andiamo ad analizzare. Viceversa nel caso del Mondrian visto prima le superfici erano adiacenti una all'altra e quindi io modificavo l'intensità luminosa ma sempre su superfici adiacenti. In questo caso abbiamo una superficie che ha questa esposizione diretta alla sorgente luminosa e un'altra superficie che invece è in una zona d'ombra, quindi il nostro confronto di riflettanze viene sempre operato, ma viene operato all'interno degli stimoli di ciascuna superficie e quindi noi percepiremo come diversi questo quadratino qui e questo quadratino qua. Sono sempre pezzettini marroni, dello stesso colore, che però appaiono di colore diverso a seconda del contesto, proprio perché il contesto su opera il confronto il SN, in un caso è dato da questi oggetti e nell'altro caso da questi oggetti. Però il meccanismo resta fisso, ed è meccanismo per cui tutti i pezzetti di questa faccia, che hanno lo stesso colore, li percepiamo sempre dello stesso colore. Padri fondatori della neurobiologia della visione dei colori: Le ricerche sono già cominciate 150 anni fa, prima che le moderne teorie cellulari fossero note. Si tratta di studi fatti senza l'interezza delle conoscenze che abbiamo adesso sul sistema visivo. La Teoria tricromatica di YoungHelmholtz è una teoria che si fonda sull'esistenza di tre sensori separati, indipendenti, uno responsabile della visione del rosso, l’altro del verde e l’altro del blu. A quel tempo non si sapeva ancora nulla di sistema visivo, quindi Helmholtz parla di tre sensori che dobbiamo avere nel nostro cervello, tre recettori diversi che ci permettono di vedere tutto l'insieme dei colori perché riescono anche a fondersi l'uno con l'altro. Quindi, dall'attività anche simultanea e integrata di questi tre sensori riusciamo a vedere tutto il repertorio dei colori. Anche la Teoria dell'opponenza cromatica di Hering-Hurvich è stata proposta in un periodo in cui si sapeva ancora molto poco del sistema visivo. Secondo questa teoria ci sono tre coppie di colori antagonisti e queste coppie sono rosso e verde, blu e giallo, e il bianco e il nero. Ciascuna di queste coppie non si fonde mai con l'altra. La differenza sostanziale tra le due teorie è che per la prima riusciamo a vedere tutti i colori anche dall'attività sincrona di questi recettori, comunque dalla fusione dei colori espressi dai diversi recettori, mentre la seconda sostiene che ci sono delle coppie che si oppongono, ma le coppie di per sé non si fondono mai perché sono antagoniste tra loro. La nostra percezione del colore in questa situazione qua deriva dal contrasto che esiste tra queste configurazioni principali. Chi ha ragione e chi ha torto? Pensiamo al sistema visivo, al sistema P (presente dalla retina fino alla corteccia), a come strutturalmente sono i nostri fotorecettori (coni e bastoncelli), come sono i pigmenti dei fotorecettori, le oxine dei recettori. Pensiamo all’importanza del contrasto. Elementi di una e dell'altra teoria si ricollegano a quanto visto sul sistema visivo. La teoria tricromatica ha una sua evidenza anatomica strutturale, proprio perché ci sono tre popolazioni di coni che hanno tre opsine diverse e queste opsine indicano la loro proprietà di assorbimento privilegiato per alcune lunghezze d'onda, e quindi di conseguenza anche di riflessione privilegiata di determinate lunghezze d'onda. Quindi, queste opsine sono sì importanti per riuscire a percepire delle differenze in termini di lunghezze d'onda, però non sono comunque di per sé ancora sufficienti ad avere la percezione cromatica, ma sono elementi strutturali che la favoriscono sicuramente. In particolare abbiamo tre popolazioni di coni che sono sensibili all'assorbimento di lunghezze d'onda differenti. Ci sono coni sensibili a lunghezze d'onda corte (quelli per i blu), coni per le lunghezze d'onda intermedie (per il verde) e quelli per il rosso sono quelli che codificano per lunghezze d'onda più lunghe. I bastoncelli hanno solo un tipo di opsina che è la rodopsina e si colloca un po’ su questa gaussiana di lunghezze d'onda, proprio perché per ciascuna popolazione di coni la distribuzione è una distribuzione gaussiana dove c'è un picco preferenziale ma c'è tutta una banda di lunghezze d'onda che riescono ad attivare in modo privilegiato quella popolazione. Quindi, i coni del rosso sono attivati per tutte le onde che rientrano nell'ambito di questa gaussiana. Ci sono intersezioni importanti tra le curve delle tre popolazioni. La teoria tricromatica si può appoggiare su questa evidenza strutturale. Un altro dato favorevole è il fatto che i difetti genetici nelle opsine causano difetti proprio nella percezione dei colori e questo è un dato clinico a sostegno del fatto che quando non funzionano le tre popolazioni di coni, quando anche solo una delle opsine viene compromessa, anche la nostra percezione cromatica è compromessa in modo significativo. I daltonici per esempio hanno un problema in una di queste opsine. Una popolazione di coni di per sé non riesce a dire nulla di più sul colore, non riesce a discriminare, ma è importante l'attività di tutte e tre le popolazioni. Questo è un dato che sembra il sostegno a favore della teoria tricromatica in cui l'autore sosteneva proprio che ci sono tre sensori e tutti e tre sono responsabili della nostra visione dei colori. Quindi se esistesse soltanto un tipo di cono, quello del rosso, noi non riusciremmo a sapere se la nostra percezione del rosso possa derivare da un’attivazione prolungata, dall’esposizione prolungata di molti fotoni di una certa lunghezza d'onda oppure soltanto all'esposizione alla sua lunghezza d'onda privilegiata, preferita. Nel senso che il fatto di avere più popolazioni di coni con opsine diverse che codificano in modo privilegiato, quindi lunghezze d'onda differenti, ci permette anche di comprendere questo fenomeno di percezione all'interno di gaussiane che hanno anche delle zone di intersezione. Quindi, si è compreso come la lunghezza d'onda preferita sia quella che corrisponde un po' al picco di attivazione, al picco della gaussiana di ciascuna popolazione, ma di base la percezione dei colori nell'ambito, ad esempio, di queste zone che sono di corrispondenza, di match, tra le popolazioni del verde e quelle del rosso in realtà dipendono strettamente dall'attività di entrambe le popolazioni dei coni e non di una sola. La teoria tricromatica non era così sbagliata perché ci sono evidenze a sostegno dell'esistenza di questi tre sensori diversi grazie ai quali vediamo tutti i colori possibili. Come vediamo tutti i colori possibili? Li vediamo grazie al contrasto, cioè grazie ad un elemento della teoria dell'opponenza cromatica. Quindi anche la teoria di Hering ha delle evidenze forti e deriva proprio dal fatto che i colori, in queste zone di overlapping, di intersezione tra le gaussiane non li percepiamo come colori miscelati, non percepiamo il rosso-verdastro o il giallo bluastro. Diamo delle considerazioni qualitativamente differenti in base al colore giallo chiaro, scuro, brillante ecc. ma, proprio il percepire come un oggetto riesca a riflettere lunghezze d'onda che appartengono a zone di gaussiana in condivisione tra due popolazioni di coni, che dovrebbero darci l'esperienza soggettiva di due colori insieme, in realtà non li percepiamo. Noi riusciamo sempre a definire un colore solo per un oggetto non ne vediamo mai due in simultanea. Quindi, di base, operiamo sempre una scelta, e questa scelta che ci permette di percepire il colore finale, la facciamo sulla base di un'operazione, di un processamento fondato sul contrasto. In base alla teoria tricromatica ci sono tre coppie antagoniste di colori che sarebbero rappresentate nella retina in tre canali differenti (tre canali nervosi distinti). Ci sarebbero delle cellule (rappresentate da questi spot centrali) che avrebbero delle zone centrali, o comunque delle zone in cui l'attivazione massima è legata a una popolazione di coni e una zona periferica in cui è massima l'attività di un'altra popolazione di coni. Parliamo di cellule essenzialmente ganglionali, quindi di cellule che ricevono l'informazione dai fotorecettori. Ci sarebbero delle cellule ganglionali che vengono eccitate massimamente quando ho uno stimolo rosso al centro e quando ho un'informazione sul verde in periferia ad esempio. E quindi cellule ganglionali (a centro on e a centro off) in cui l'attivazione è legata però non soltanto alla posizione dello stimolo ma anche al colore dello stimolo. Questa sarebbe un’ulteriore attività delle cellule ganglionali, non di tutte, di quelle del sistema P. Ricordarsi la suddivisione in sistema M e in sistema P: solo le parvocellulari saranno strutturate in questo modo, quindi saranno quelle che codificano anche per l'informazione sul colore e lo fanno sempre sulla base di un contrasto. Il contrasto, in questo esempio, è un contrasto dato da un segnale che ha un’informazione rossa nel centro e un'informazione verde nella periferia o, in questo caso, un segnale che ha un'informazione blu nel centro e un'informazione gialla in periferia, dove il giallo è legato proprio all'attività simultanea di queste due popolazioni di coni, del rosso e del verde. La Teoria dell’opponenza cromatica di Hering diceva che ci sono coppie antagoniste, una prima coppia è quella rossa-verde e un'altra coppia è blu-giallo, dove il giallo viene in realtà codificato da queste due popolazioni di coni insieme, perché non abbiamo dei coni specifici per il giallo, e il giallo rientra nell'ambito di gaussiane che rispondono sia alle lunghezze d'onda codificate dai coni del verde che quelle codificate dai coni rossi. Queste cellule sono cellule ganglionali nella retina. Cellule di questo tipo ci sono anche a livello della stazione successiva, del corpo genicolato laterale, quindi nel talamo. Quindi, cellule ganglionali retiniche opponenti per il rosso e il verde, per il blu e il giallo e anche per il bianco e nero (il chiaro e lo scuro). Non c'è soltanto un tipo di cellula ma almeno due tipi di cellule opponenti: le singole opponenti e le doppie opponenti. Una cellula opponente è una cellula che riesce a codificare per l'informazione del contrasto sul colore in zone diverse del suo campo percettivo. Le cellule opponenti, cellule sensibili al contrasto di colori, generalmente sono le cellule ganglionali nella retina. Però possono essere opponenti anche le cellule talamiche del corpo genicolato laterale e le cellule della corteccia nell'area blob di V1 o anche nelle aree di corteccia extra striata. La corteccia visiva primaria, la corteccia striata, chiamata V1 è soltanto una delle cortecce visive, ma ci sono anche aree extra striate importanti, ad esempio V4, che è una regione di cellule doppie opponenti, è una delle aree fondamentali per la visione dei colori. Queste cellule si trovano anche ai livelli superiori, anche a livello della corteccia, proprio perché il sistema P si mantiene in tutti gli stadi. Sono le cellule che già dalla retina hanno questa proprietà di afferire al sistema parvo cellulare, che è responsabile di elaborare l'informazione sul colore, queste proiettano a cellule del sistema P a livello del talamo, e la stessa cosa è mantenuta in corteccia, dove le regioni deputate all'analisi del colore sono le regioni blob (quindi il terzo sistema di colonne di un ipercolonna). Il filo di tutto il sistema P poi converge sulle colonne blob della corteccia striata. Quando si dice cellula opponente, senza la specifica di che tipo di cellula sia, occorre pensare in primo luogo alle cellule ganglionali però bisogna sapere che le cellule opponenti ci sono a tutti i livelli. Differenza tra le singole opponenti e le doppie opponenti Le doppie opponenti hanno un campo recettivo doppio. La singola opponente ha una zona centrale di campo recettivo, che è sensibile alla luce rossa, quindi se in questa zona si presenta uno stimolo che attiva la popolazione di coni che codificano per il rosso, avrò un aumento della frequenza di scarica, però poi questa stessa cellula è anche sensibile alla luce verde nella zona periferica, e quindi l'informazione del verde in periferia è un'informazione che inibisce l'attività della stessa cellula. L'attività finale è un'attività che terrà conto sia dell'eccitazione del centro che dell'inibizione della periferia, quindi del contrasto tra centro e periferia. Un'eccitazione al centro causata dal rosso produce un aumento di scarica di questa cellula, però un’inibizione sul verde produce una diminuzione della frequenza di scarica. Quindi, se ci fosse un'opposizione forte, se lo spot fosse idealmente un pallino rosso circondato da un fascio verde, in realtà non si ha attività perché le zone eccitatorie e inibitorie si elidono. Però, sono cellule che codificano sempre per le differenze che esistono dell'eccitazione rossa al centro e verde in periferia. Questo è solo un esempio di opposizione di colori ma ci sono cellule opponenti per tutte le coppie antagoniste, quindi quelle che si eccitano quando c'è il blu al centro e il giallo in periferia, quando c'è il giallo al centro e il blu in periferia, quando c'è il bianco e il nero, quindi comunque le cellule singole e doppie opponenti rispondono al contrasto tra le diverse coppie antagoniste. Le doppie opponenti sono leggermente più complesse come struttura perché rispondono al contrasto che c'è tra aree adiacenti, nel senso che il contrasto è sempre all'interno della stessa zona, rilevano il contrasto nella zona centrale e nella zona periferica, ma sempre facendo una distinzione tra centro e periferia. Occorre immaginare che anche quella porzione centrale di campo recettivo in realtà sia codificata come differenza di contrasto tra la parte più centrale e la parte più periferica della zona centrale, e lo stesso in periferia, percepiscono il contrasto tra la porzione più centrale e quella più periferica della stessa periferia. Il campo recettivo è come se si sdoppiasse perché diventano due campi per il campo centrale e due campi per il campo periferico, perché hanno zone eccitatorie e inibitorie sia al centro che in periferia. Di fatto l'informazione sul contrasto è ulteriormente rafforzata perché riesce ad essere rilevata in ciascuna delle parti, sia nelle porzioni centrali che nelle porzioni periferiche. Queste sono le cellule prevalentemente localizzate nell'area V4 che è l'area di corteccia extrastriata che si occupa in modo privilegiato dell'esame dell'informazione sul colore. Le doppie opponenti sono proprio nell’area privilegiata per i colori, quindi rafforziamo dal punto di vista anatomico le cellule specializzate per quella funzione. Mentre in V1, dove comunque arrivano tutte le informazioni, anche quelle del sistema magno cellulare, ho delle cellule che fanno operazioni di analisi di contrasto più grossolane o comunque non così precise. In sintesi: Due teorie storiche: teoria tricromatica e teoria dell'opponenza cromatica. La teoria tricromatica ha un dato anatomico a suo favore oltre che un dato clinico: il dato anatomico è che effettivamente ci sono tre popolazioni di coni con opsine differenti, spettri di emissioni e assorbimento preferenziale per determinate lunghezze d'onda. A livello clinico difetti in una popolazione delle opsine hanno effetti diretti sulla nostra visione dei colori. Quindi, la teoria tricromatica sembra essere supportata. Nello stesso tempo sono presenti elementi della teoria dell'opponenza cromatica perché comunque il contrasto tra coppie antagoniste di colori è l'informazione più saliente per la nostra codifica della percezione visiva, proprio perché esistono cellule ganglionali del sistema P parvocellulare che già si presentano come cellule opponenti, che quindi sono più stimolate quando ho un'informazione di centro e periferia, più sensibili al contrasto tra coppie di colori antagonisti. Queste cellule opponenti continuano ad essere presenti negli stadi successivi di elaborazione visiva, le troviamo nel corpo genicolato laterale del talamo, le troviamo nella corteccia visiva sia primaria V1 che extra striata (le aree visive secondarie). In particolare a questi livelli la complessità aumenta ulteriormente perché abbiamo anche cellule doppie opponenti che hanno questa informazione sul contrasto in tutta la porzione del campo recettivo, nel senso che discriminano il contrasto sia nel centro che nella periferia del campo recettivo, ma questo contrasto è sempre fondato sulle coppie di colori antagoniste fra loro, quindi su ciò che era stato teorizzato nella teoria dell'opponenza cromatica. Anche molto prima delle teorie della neurobiologia sulla visione c'era chi è arrivato sperimentalmente a comprendere l'importanza del contrasto tra coppie di colori opponenti. Anche noi sperimentalmente possiamo provare delle sensazioni di colori fondate sull'analisi del contrasto: Se ci si concentra su uno dei quattro stimoli presentati e poi si fissa lo stimolo centrale in modo prolungato, la percezione del colore dei quattro quadratini risulta essere modificata dai colori visti in precedenza. Una configurazione stimolo con tanti ritagli sotto sorgenti luminose di tipo differente, lo sfondo globale ci dà informazione sul contrasto perché un ambiente completamente giallo o un ambiente completamente blu ci fa virare il colore di un oggetto verso il colore complementare che è dato dallo sfondo. In questo caso la percezione errata è basata sul contrasto cromatico simultaneo. Percezione della distanza Quando voglio avere un'informazione sulla distanza di un oggetto da me, utilizzo meccanismi diversi a seconda del tipo di distanza che esiste rispetto a un punto di fissazione. Ho un punto di fissazione (b) che in questo caso è frontale a me, questo punto viene codificato in due porzioni di retina dette omonime, perché sono proprio punti corrispondenti delle nelle due retine. Le informazioni sulla distanza degli oggetti dal punto di fissazione, e quindi anche l'informazione di profondità e di tridimensionalità, che si basa sul concetto di distanza si ottiene in un altro modo cioè si ottiene grazie alla disparità binoculare. Il fenomeno di disparità binoculare viene elaborato fino a livello corticale, quindi si tratta di stimoli che vengono proiettati, in realtà, su regioni diverse delle due retine ed è proprio la distanza rispetto al punto di fissazione che viene interpretata dal mio sistema nervoso come profondità. Ci sono proprio neuroni corticali che si attivano solo in risposta a queste disparità retiniche lontane o vicine. Quindi, nella corteccia visiva striata ed extra striata, in particolare, avrò una popolazione di neuroni che si attiva quando voglio considerare i punti che sono o lontani o vicini rispetto al mio punto di fissazione, e mi fa percepire questi punti come più lontani o più vicini. Il meccanismo è proprio considerare l'immagine che arriva in modo diverso già nelle due retine. Tuttavia, questo processo di codifica della distanza dal punto di fissazione in realtà non vale sempre, vale soltanto per distanze limite, fino a 10 m. Oltre i 10 m si attiva la cosiddetta la visione ciclopica nel senso che la mia percezione visione della distanza, della profondità, sarà basata su considerazioni di tipo diverso, che sono chiamate indizi pittorici di profondità. Di base accade che la mia corteccia riesce già a elaborare l'informazione sulla distanza, e quindi anche sulla profondità degli oggetti, se gli stimoli sono entro i 10 m, facendo proprio un calcolo che si basa sulla disparità binoculare, quindi su zone diverse di retina in cui arrivano le immagini rispetto al punto di fissazione. C'è proprio un calcolo attivo della distanza dello stimolo rispetto al punto di fissazione di quel momento. Punto di fissazione o piano di fissazione perché si parla di tutto un livello su cui il soggetto mantiene lo sguardo, e nello stesso tempo riesce a vedere sia ciò che è vicino, sia ciò che è lontano facendo proprio questo calcolo della distanza, distanza che c'è già a livello della retina perché i punti hanno distanze diverse già a livello della retina. Viceversa, per distanze superiori ai 10 m, abbiamo gli indizi pittorici di profondità. Principio di familiarità: dice che noi consideriamo gli oggetti a distanze superiori ai 10 m (con visione ciclopica) più lontani o più vicini sulla base delle loro dimensioni. Quindi quelli più vicini sono quelli che ci sembrano più grandi, quelli più piccoli sono quelli più lontani. Interposizione: dice che se vedo un oggetto che è parzialmente coperto da un altro, percepirò quello che viene coperto come quello più lontano. L'oggetto che lo copre è l'oggetto più vicino a me. Prospettiva lineare: secondo cui quando vi sono due rette parallele io le percepisco come realtà convergenti a mano a mano che sono lontane da me, dal mio punto di vista. Più due rette parallele sono lontane più tendo a percepirle come convergenti. Trasparenza: principio in base al quale le cose che vengono percepite attraverso un buco, una fessura sembrano più lontane. Effetto di parallasse: in base al quale percepiamo come più vicini gli oggetti che si muovono più velocemente rispetto ad oggetti che si muovono più lentamente. Quelli più lenti sono percepiti come più lontani. Questi sono indizi che aiutano la nostra ricostruzione della distanza, della profondità e della tridimensionalità di una scena visiva. Quindi, ci sono meccanismi diversi per distanze inferiori o maggiori rispetto al soggetto. La corteccia visiva occupa di base un terzo dell'intera corteccia cerebrale e quindi non parliamo soltanto della corteccia visiva primaria del lobo occipitale, ma di tutte le sottoaree che si distribuiscono nei settori anche parietali e temporali. Si tratta di sottoaree fondamentali per operare questa analisi parallela delle caratteristiche diverse dello stimolo, quindi delle submodalità diverse della visione, la forma, colore e il movimento. Quando abbiamo esaminato in dettaglio come funzionano le cellule semplici e complesse della V1, abbiamo parlato essenzialmente di analisi della forma, che si basa sull'analisi dei contorni, dei bordi delle superfici perché ci sono cellule in grado già in V1 di scomporre tutti gli elementi, quasi punto a punto, del contorno di un oggetto e poi andarlo quasi a ricostruire, nel caso delle cellule complesse, nella sua struttura globale. Tuttavia ci sono anche delle ulteriori aree extra striate, che nel caso della forma corrispondono all’area V3, nel caso del colore a V4 e nel caso del movimento a V5, che sono responsabili dell’ analisi parallela delle informazioni caratteristiche di ogni singola submodalità. Quindi, l'analisi della forma, di per sé, è un esempio eccezionale di analisi gerarchica nel sistema visivo, però l'analisi globale di uno stimolo visivo si fonda comunque su un’analisi parallela, che è parallelamente condotta in regioni extra striate specializzate, ma l'informazione di base comunque converge primariamente in V1. Quindi, nell'area visiva primaria V1 arrivano comunque tutte le informazioni che abbiamo visto fino a questo momento. C'è un'area V2 che è organizzata allo stesso modo di V1 e si trova nelle porzioni occipitali limitrofe, che quindi ha lo stesso tipo di struttura di ipercolonne, quelle di dominanza oculare, quelle di orientamento, le regioni blob e interblob. Poi però le connessioni da V1 e V2 vanno in modo privilegiato alle diverse sottoaree a seconda del tipo di analisi che si deve portare avanti. Le regioni blob andranno a proiettare essenzialmente in V4. Le regioni interblob, che sono invece quelle porzioni delle ipercolonne specializzate nella forma e nel movimento, proietteranno a V3 e V5. Tutte queste sotto aree sono sia nei lobi parietali che nei lobi temporali. Una evidenza dell'esistenza di queste sottoaree è legata anche alla clinica, nel senso che sono descritti pazienti con lesioni selettive e disturbi specifici delle submodalità visive. Ad esempio si è visto come lesioni a livello di V3 hanno conseguenze dirette sulla nostra percezione delle forme degli oggetti, pazienti con lesioni in V4 hanno la cosiddetta acromatopsia come conseguenza diretta per cui riescono a vedere i colori nelle proprietà base, le proprietà che arrivano a livello di codifica fino a V1, V2 ma perdendo l'attività di V4 si perde tantissimo della percezione dei colori. Fino ad oggi è stata descritta in letteratura una sola paziente che presenta lesione specifica a livello V5 e quindi sintomi chiamati akinetopsia, quindi non vede gli oggetti che si muovono. Questa è specifica per gli stimoli visivi. Un'altra prova dell'esistenza funzionale di sotto aree è data dagli esperimenti di inattivazione temporanea e selettiva dei neuroni di queste aree. Si è visto che silenziando l'attività di queste regioni, di fatto, si produce il deficit corrispondente alla funzione esercitata, quindi ci sono prove forti dell'esistenza di questi meccanismi paralleli di analisi delle sub modalità visive. Nota importante sull'esistenza di due tipi di connessioni principali di V1: V1 e V2 raccolgono tutta questa informazione vista fino ad oggi, la trasmettono poi alle aree secondarie extrastriate V3, V4 e V5, ma ci sono due circuiti privilegiati, che sono il circuito V1-V4 e il circuito V1-V5. Questo esempio di configurazioni stimolo serve per farvi prestare attenzione a questi due cerchi centrali, che ci appaiono percettivamente come di dimensioni diverse, proprio perché li confrontiamo. Li percepiamo sulla base delle differenze con i cerchi che li circondano. Di fatto però, se togliamo queste differenze, vediamo come di base non c'è differenza tra le dimensioni di questi due cerchi. Quindi, se dovessimo (con le dita della mano) andare ad afferrare questo cerchio centrale o quest'altro cerchio, si potrebbe pensare che andremmo a mettere le dita a distanze diverse per prendere l'oggetto che ci sembra più grande e quello che sembra più piccolo, ma in realtà non è così, perché le nostre mani, le nostre dita, e quindi la nostra corteccia codifica già l'informazione corretta sulla dimensione di quell'oggetto. E questo è possibile perché questi sistemi paralleli sono simultaneamente operativi e ci sono 2 sistemi importantissimi che permettono di codificare una rappresentazione diversa di quest'oggetto, una rappresentazione del cosa e una rappresentazione del dove; quindi abbiamo già un'immagine di che cosa è questo oggetto, che ci fa sembrare un cerchio più grande e l'altro più piccolo, ma c'è già anche un sistema attivo di informazioni e rappresentazioni che codifica sul dove, sulla posizione e che mi fa aprire le dita della mano proprio sulla base della posizione, non tanto sulla base della dimensione percepita. V1 e V2 sono a livello di corteccia occipitale e poi V4 e V5. Il sistema V1,V2,V4, è il sistema di connessione con la corteccia infero temporale ed è fondamentale per la via del cosa, la via del riconoscimento degli oggetti. Viceversa, le connessioni V1, V2 con V5 che è importante per l'analisi del movimento ma è anche importante perché queste connessioni con la corteccia parietale posteriore sono fondamentali per l'analisi della posizione spaziale dell'oggetto, quindi del dove. Queste due vie sono contemporaneamente attive e si correggono anche una con l'altra, come nel caso dell'oggetto che devo andare a prendere, ho una rappresentazione che mi dà informazioni sul cosa ma ho anche un'informazione privilegiata sulla posizione che coordina il mio movimento. V5 è importante sui movimenti, di fatto i neuroni di V5 codificano per dei vettori che indicano il movimento, la direzione del movimento dell'oggetto, quindi in questo caso dello stimolo visivo, e quindi popolazioni che esprimono dei vettori di direzioni differenti. Un punto ancora aperto nella ricerca è come facciamo in realtà di integrare tutto questo? Come facciamo ad avere un'informazione dello stimolo che sia un'informazione in contemporanea efficace di tutte le sue caratteristiche? I processi neuro anatomici e fisiologici con cui, di fatto, si ha l'integrazione, sono ancora oggetto di studio attuale della ricerca. 16. Sistema uditivo Il suono è una compressione e rarefazione di molecole d'aria proprio perchè il mezzo preferenziale delle onde acustiche. Il suono è dato dalla vibrazione di un mezzo elastico che si propaga con un modo ondulatorio e rettilinio. C'è una sorgente di emissione motoria, che vibra e le vibrazioni si trasmettono nell'aria e causano una progressiva compressione e rarefazione delle molecole del mezzo, che si traduce in onde pressorie. La velocità di propagazione sarà dipendente dal mezzo. Sarà 13040 m/s quando il mezzo è l'acqua. Le due caratteristiche essenziali del suono sono la frequenza e l'ampiezza. La frequenza indica il numero di cicli al secondo ed esprime il concetto di altezza di un suono (acuto o basso) e l'unità di misura della frequenza è l'Hz. Le frequenze si distribuiscono tra 20 e 20000 Hz. Un'altra caratteristica è l'ampiezza, correlata con l'intensità della stimolazione: ci dice se un suono è forte o debole e l'unità di misura è dB. La maggior parte dei suoni è un insieme di onde diverse. Abbiamo tre tipologie di suoni: suoni puri, che per poterli sentire devono essere riprodotti in modo artificiale e hanno una sola frequenza caratteristica. La maggior parte dei suoni è composto da onde diverse, cioè la sorgente emette per più frequenze caratteristiche. Se vogliamo rappresentare queste onde, si ha sempre una frequenza fondamentale da cui derivano le armoniche. Ci sono i rumori, viceversa, che non sono rappresentabili in onde perchè non hanno la periodicità che caratterizza l'onda. Parliamo di misure sia fisiche che psicofisiche quando trattiamo i suoni. Le misure fisiche sono la frequenza e l'intensità. Misurare l'intensità del suono significa misurare la variazioni di pressione. Farlo non è semplice e nella pratica in ogni suono si fa un rapporto tra una pressione di riferimento (pressione minima necessaria per ottenere una sensazione uditiva) con la pressione dello stimolo da misurare. Le misure psicofisiche richiamo i concetti di soglia. Si tratta delle possibilità che si hanno sperimentalmente di misurare la capacità di ogni soggetto di discriminare stimoli. La soglia di udibilità si distingue in soglia di discriminazione in intensità e soglia di discriminazione in frequenza. La capacità di discriminare è migliore nelle frequenze intermedie e peggiora quando si passa a frequenze più alte o più basse. Ci sono delle curve, che si chiamano isofone, che hanno come unità di misura il Phon. Questa curva esprime l'intensità di ogni data frequenza. Questa però non ci dice nulla sulla sensazione acustica soggettiva, ma ci dice soltanto che per quella frequenza ci possono essere intensità diverse. Non si tratta di parametri psicofisici dunque. Per far questo si usa come unità di misura il Sone, che è l'intensità soggettivamente percepita di un suono che ha frequenza costante. Iniziamo ad analizzare l'organo principale per la ricezione dei segnali acustici. Le suddivisioni principale riguardano l'orecchio esterno, medio e interno, che intervengono in modo funzionalmente diverso nell'elaborazione dello stimolo acustico. Quando parliamo di orecchio esterno si fa riferimento alla pinna, che delimita il padiglione auricolare, continua col meato uditivo esterno, e termina con l'inizio dell'orecchio medio che si ha con il timpano. La funzione principale è quella di raccogliere l'energia sonora e convogliarla sul timpano. La pressione si ottiene con il rapporto tra frequenza e superficie, quindi in questo caso la possibilità di aumentare la pressione attraverso la superficie è notevolmente sfruttata sia dalla conformazione della pinna che dal canale del meato acustico. Un'altra funzione importante è quella filtrare le frequenze, cioè sulla base della provenienza dello stimolo sonoro, la conformazione dell'orecchio esterno facilita il processo di localizzazione. L'orecchio esterno principalmente fa confluire l'onda acustica verso la membrana timpanica. L'orecchio medio è costituito dalla membrana timpanica e da tre ossicini diversi, che sono martello, incudine e staffa, che sono una specie di sistema di leve connesse una all'altra. La staffa poggia su una finestra ovale. La funzione dell'orecchio medio è quella di riuscire a superare l'impedenza. Una volta che il suono passa dall'aria ai liquidi interni della coclea, cambiando mezzo di propagazione, quindi serve un sistema che faccia superare questa impedenza elevata dei liquidi. Serve, per farlo, un aumento della pressione. Tutto questo viene svolto dalla superficie del timpano, che è molto grande rispetto al diametro della finestra ovale, e grazie al sistema di leve dato da questi tre ossicini. A questo punto le vibrazioni entra nell'orecchio interno tramite la finestra ovale. Essenzialmente parliamo di coclea, che è la porzione ricettoriale vera e propria. Ha una forma carattestica a chiocciola. È larga 10 mm e immaginando di allungarla lunghezza totale è di 35 mm. La composizione interna della coclea è fatta a compartimenti, che contengono i liquidi. Si parla di scala vestibolare (che contiene perilinfa), scala media (endolinfa) e scala timpanica (perilinfa). Tra scala media e timpanica c'è una membrana basilare, che ha una porzione superiore chiamata membrana tettoria. Questo sistema di membrane rappresenta l'organo recettoriale del Corti che rappresenta il vero e proprio sistema recettivo. L'organo del Corti è importante perchè, oltre al fatto che qui avviene la trasduzione del segnale, svolge un'analisi spettrale del suono, cioè lo scompone nelle sue frequenze in due stadi: prerecettoriale (membrana basilare) e recettoriale (cellule ciliate). Le cellule cilate interne o esterne hanno una porzione ancorata alla membrana che non fuoriesce e un ciuffo di ciglia, disposte a pettine, che fuoriescono dal liquido ed entrano in contatto con l'endolinfa della scala media. Questi recettori sono di II ordine, che rappresenta un elemento cellulare indipendente e autonomo che riceve lo stimolo, in questo caso la vibrazione dalla finestra ovale, e lo trasduce in segnale elettrico e le invia nel ganglio spinale. Nella trasduzione del segnale la finestra rotonda, subito dopo la finestra ovale, svolge un ruolo altrettanto importante. Quando arriva un onde, la vibrazione passa attraverso la catena di ossicini e la finestra ovale e causa uno spostamento del liquido nella coclea. Questo fa sì che le due parti dell'organo del Corti, cioè le due membrane, vibrino in direzioni opposte. Questo movimento fa sì che le ciglia delle cellule ciliate viene continuamente stirato da un lato e dall'altro, quindi si ha uno stiramento meccanico causato dall'arrivo della vibrazione. C'è dunque una differenza pressoria tra scala vestibolare e scala timpanica che viene tramessa alla scala media che vibra. Da questa descrizione si può dedurre che si parla non solo di recettori di II ordine, ma meccanocettori. L'endolinfa in condizione di riposo è ricco di ioni potassio, quindi ha un potenziale intorno a +80 mV. Il liquido intracellulare contenuto nel compartimento della cellula recettrice ha un potenziale di membrana intorno a -60 mV. Tra i due liquidi la differenza di potenziale è quindi molto alta. Ogni ciglio è collegato al ciglio successivo tramite un filamento detto tip-link, che ha la funzione di apertura di canali. Ci sono canali che si aprono meccanicamento a seguito di stiramento del tip-link. In condizioni di attivazione, quando arriva l'onda pressoria, che causa spostamento meccanico delle membrane e lo stiramento del tiplink nella sua estensione massima. Lo stiramento del filamento fa aprire i canali situati sulla punta dello stereociglio, che permettono l'ingresso del potassio K all'interno della cellula recettrice. Il risultato finale è una depolarizzazione della cellula. Questo è l'inizio del meccanismo di trasduzione. Oltre a depolarizzazione, l'apertura dei canali causa anche apertura dei canali del calcio, con ingresso di calcio nella cellula. Il risultato finale di questo processo è un'ulteriore depolarizzazione e rilascio di neurotrasmettitore. Quando lo stimolo continua nel tempo un ulteriore fenomeno è quello dell'adattamento allo stimolo: una componente proteica sullo stereociglio (miosina 1β) scivola lungo il citoscheletreo di actina spostando verso il basso il punto di inserzione del tip-link sullo stereociglio e quindi la vibrazione non è sufficiente a produrre lo stiramento necessario a produrre il canale. È un processo che spiega come mai non recepiamo tutti i suoni. I punti A, B, C e D rappresentano punti diversi dell'attivazione delle cellule recettrici. Il punto A e D rappresentano le condizioni iniziali, di riposo, in cui la differenza di potenziale è di -60 mV, mentre l'endolinfa è ricca di potassio. L'onda pressoria fa sì che i liquidi all'interno della coclea si spostano, si muovono le membrane, si tirano le ciglia in direzione dei filamenti più alti; questo stiramento, grazie al tip-link, che connette un ciglio al successivo ed è collegato al canale del potassio, si apre la porta di questi canali; il potassio entra nella cellula e causa una prima depolarizzazione, che fa cambiare le differenze di potenziale e comporta altri due fenomeni: apertura dei canali per il calcio voltaggio-dipendenti causando l'entrata di ioni calcio con un'ulteriore depolarizzazione. Tutto questo è ciò che permette di raggiungere il potenziale di recettore, causando rilascio di neurotrasmettitore. È un fenomeno che si risolve in tempi brevissimi. Ci sono stimoli sonori che non attivano questo processo o causano un'adattamento. La membrana basilare non vibra allo stesso modo per tutte le frequenze. Nelle alte frequenze vibra la base, per le basse l'apice. Essa è più larga è flessibile all'estremità apicale e più stretta e rigida alla base. Ciò facilita il fatto che ogni punto vibra di più in risposta ad una certa frequenza con conseguente rappresentazione topografica delle frequenze (tonotopica). Ciò che è stato studiato da Helmotz. Fino ad allora si riteneva che il processo di analisi acustica fosse passivo. Anche le cellule recettrici rispondono in modo differente alla frequente: si è visto con esperimenti specifici che cellule ciliate diverse rispondono in modo preferenziale a frequenze diverse. Avvicinando un microfono alla coclea, si è visto che essa stessa emette suoni, scoprendo così che non è soltanto un recettore. Essa tessa in modo attivo inizia a vibrare con suoni che hanno stessa frequenza di quelli ricevuti. A bassi livelli di intensità sonora c'è anche quindi un processo attivo: cellule ciliate esterne ricevono efferenze da complesso. La frequenza caratteristica è codificata dunque sia dalla membrana basilare sia dalle cellule ciliate, in cui un sottogruppo vibra con la frequenza dello stimolo ricevuto, che aumentano l'ampiezza e restringono l'estensione del picco di vibrazione. Le ciliate interne sono circa 3500 e quelle esterne sono circa 12000, sia perchè le esterne sono disposte in tre file, con un totale di 16000. I neuroni dei gangli sono molti di più. Le connessioni sono da un lato divergenti e dall'altro convergenti, nel senso che ciascuna cellula ciliata interna va a contattare almeno 10 neuroni diversi del ganglio di Corti. Viceversa, per le cellule ciliate esterne ci sono connessioni convergenti. Ciascun neurone del ganglio di Corti riceve da più cellule esterne. Quando è massimo il grado di divergenza, sarà aumentata molto la capacità di discriminazione. Quindi i recettori veri e propri, che discriminano molto, sono le ciliate interne. Vediamo le vie acustiche centrali. Innanzitutti abbiamo i neuroni sensitivi primari nel ganglio del Corti; le loro terminazioni arrivano a livello dei nuclei cocleari, che si trovano nel tronco. Da qui la via diventa crociata a livello dell'oliva superiroe. Accanto a questi, ci sono i nuclei vestibolari, che ricevono dal labirinto vestibolare. Abbiamo tre stazioni principali a questo punto: nucleo del lemnisco laterale, collicolo inferiore, nucleo genicolato mediale del talamo. Il collicolo inferiore è quella stazione che facilita la reazione di orientamento degli stimoli uditivi. Importante è la rappresentazione tonotopica, che codica per le frequenze degli stimoli, che è presente in tutte le stazioni centrali, dalla coclea alla corteccia uditiva primaria, a livello del lobo temporale. La terminologia con cui la si indica è A1. La localizzazione del suono è mediata da processi che riescono a codificare per differenza di intensità e frequenza. Sotto i 3000 Hz vengono decifrate le differenze di tempo tra un orecchio e l'altro. Sopra i 3000 Hz la codifica si basa su differenze di intensità, in quanto .la testa agisce da ostacolo, quindi il suono arriva più debole dall'altro orecchio). Le differenze di tempo interneurali sono minime, anche di 10 nanosec. Ci sono neuroni (che hanno come stazione l'oliva superiore mediale) che si attivano solo quando gli stimoli sono perfettamente sincroni, mentre sono inattivi quando la differenza è anche di 10 nanosec. La differenza di intensità è codificata da fibre che passano dall'oliva superiore laterale. Quando lo stimolo proviene dalla porzione di sinistra arriva di più all'orecchio ipsilaterale: ci sono un gruppo di neuroni che si eccita maggiormente aumentando la frequenza di scarica. Il nucleo del lemnisco laterale è associata all'inizio del suono e la sua durata. Il collicolo inferiore media l'orientamento ma non è stata trovata una vera e propria mappa uditiva. Come negli altri sistemi, anche qui l'informazione è distorta. Una caratteristica interessante è che la corteccia uditiva primaria non corrisponde all'area di Wernicke. Questo indica che è un pezzo di corteccia che serve a decodificare i suoni elementari che possono poi essere ulteriormente codificate. La rappresentazione finale è in ipercolonne: da un lato ci sono le colonne che codificano le frequenze degli stimoli, dall'altro quelle che codificano la possibilità di attivazione per stimoli che provengono da entrambe le orecchie o che provengono da un solo orecchio. Ci sono colonne di cellule EE (neuroni eccitati da entrambe le orecchie) ed EI (neuroni eccitati da un orecchio e inibiti dall'altro). 17. Olfatto e gusto L'aria entra tramite la cavità nasale e incontra una serie di neuroni detti neuroni olfattivi che sono bipolari, che si trovano su tutto l'epitelio olfattivo, strato epiteliale che riveste la superficie interna del naso. I neuroni sono bipolari, quindi hanno un assone e un solo dendrite. L'assone va verso il bulbo olfattivo, il dendrite è ancorato all'epitelio olfattivo. L'uomo non ha un olfatto ben sviluppato; abbiamo 12 milioni, mentre il cane 1 miliardo. I recettori olfattivi si rinnovano ogni 60 giorni, rigenerano continuamente il loro assone e anche i neuroni del bulbo olfattivo sono continuamente riprodotti e sostituiti nel corso della vita. I recettori terminano nelle ciglia olfattive, porzione recettrice vera e propria in grado di agganciare le molecole recettrici. Le ciglia sono avvolte da uno strato muco, che ha funzione di protezione e di controllo dell'ambiente ionico delle ciglia. L'esperienza comune è che quando questo strato di muco si ispessisce la capacità olfattivo è notevolmente ridotta perchè le ciglia riescono peggio a legarsi alle diverse sostante chimiche. La sostanza odorosa si lega al recettore specifico rappresentata dalle cigle olfattive; da quest'interazione ligando-recettore si avvia il processo di trasduzione del segnale, che avviene attraverso i soliti meccanismi ionotropi e metabotropi. Ciascun recettore olfattivo è specializzato per avere siti di legame specifici per le diverse sostanze. Un altro elemento sono le chiangole di Bowman che hanno la funzione di produrre il muco. L'assone di questi recettori converge nel bulbo olfattivo attraverso il nervo olfattivo. Dal bulbo olfattivo le proiezioni dei neuroni andranno tramite il tratto olfattivo direttamente a centri corticali. Si tratta di un sistema atipico perchè non ha la stazione talamica. Il talamo interviene in un secondo tempo, ma il primo tipo di connessione che fuoriesce è diretto a centri corticali. L'arrivo del recettore attiva la proteina G che attiva a sua volta una serie di reazioni a cascata e infine l'apertura di ioni sodio. Si ha una depolarizzazione e potenziale d'azione. I recettori olfattivi stessi riescono a generare potenziali d'azione a differenza di altri recettori sensoriali. Centinaia di proteine recettrici diverse sono specifiche per uno pochi odoranti. Ogni cellule esprime un piccolo set di tali proteine ed è quindi sensibile solo a certe sostanze odorose. Tanto più la sostanza è concentrata tanto più riuscirò ad attivare recettori che hanno proteine specifiche in grado di riconoscere le sostante. Le ricerche hanno cercato di classificare le sostanze odorosa in base alla struttura molecolare della sostanza. Si è visto che la qualità stessa degli odori può cambiare con la concentrazione e che riconosciamo come odori diversi anche due molecole che sono due isoforme della stessa molecola. È stato studiato nel roditore la convergenza di assoni di neuroni olfattivi che hanno la stessa proteina recettrice: 2500 assoni di recettori olfattivi convergono circa su circa 25 cellule mitrali. Anche a livello del bulbo olfattivo, come abbiamo detto, c'è un continuo rinnovo cellulare. Le cellule periglomerulari e i granuli si rigenenano tutta la vita a partire da cellule staminali: ciò avviene nella zona sottoventricolare. Qui c'è una popolazione che matura e va incontro a tre diversi stadi di maturazione. Sono gli unici neuroni del nostro cervello che possono continuamente ricrearsi. Nei recettori olfattivi, attraverso il nervo olfattivo, si inviano le informazioni al bulbo olfattivo. Da qui le fibre si raccolgono nel tratto olfattivo e vanno a contatto diretto con la corteccia periforme. Da qui ci sono ulteriori connessioni che vanno alla corteccia orbitofrontale e strutture connesse con il talamo. La corteccia orbitofrontale media la percezione cosciente degli odori nell'uomo. Altre funzione importanti degli stimoli olfattivi come quelle di dare risposte vegetative ed emotive, importanti per i comportamenti riproduttivo, alimentare e aggressivo si ottengono grazie alle connessioni con le stazioni ipotalamiche. La formazione ippocampale è invece importante per le componenti affettive dell'olfatto e memoria. Quello che non è chiaro però è come è rappresentata la stimolazione olfattiva a livello corticale, come non è chiaro rispetto all'udito una rappresentazione corticale dello spazio acustico esterno. Le popolazioni di recettori per il gusto sono ristrette e localizzati nelle pupille gustative. Qui ci sono i bottoni gustativi che rappresentano gli elementi laterali della papilla gustativa. All'interno ci sono i recettori specializzati per legarsi allo stimolo. I bottoni gustativi si trovano sulla lingua, sul palato, sulla faringe, etc, e sono responsabili di elaborare informazioni sull'identità, sulla concentrazione e sulla qualità (sgradevole, gradevole, nocivo) delle sostanze chimiche contenute nel cibo. Queste informazioni preparano anche l'apparato gatrointestinale a ricevere e digerire il cibo (attivando la salivazione e la degustazione). L'informazione relativa al dolce, ad esempio, viene elaborata a livello dei bottoni che si trovano sulla punta, mentre quella al salito avviene grazie alle porzioni laterali e superiori della lingua, invece sull'acido porzioni laterali e posteriori. La porzione posteriore è importante anche per l'amaro. Mentre le informazioni relative alla temperatura e composizione del cibo (viscosità) sono trasdotte dai recettori somatosensoriali del nervo trigemino e altri nervi cranici e trasmesse dalla lingua e dalla bocca al talamo e alle cortecce somatosensoriali. La soglia di concentrazione per sostanze non nocive è alta, nel senso che riusciamo ad avere una soglia minima per riuscire a discriminare il gusto di una sostanza con una concentrazione alta di quella sostanza, mentre per quelle parzialmente dannose è bassa. Le soglie di sensibilità sono diverse nelle diverse zone della lingua. Le informazioni elaborate da ciascuna linea di recettori rimangono separati anche a livello corticale (si parla di codice delal linea attivata). Esperimenti di imaging cerebrale dopo assunzione di determiante sostanze con gusti diversi hanno permesso di vedere le sedi cerebrali leggermente diverse per ciascun tipo di stimolo, dimostrazione del fatto che si ha un'elaborazione per vie parallele indipendenti. Gli stimoli dati dalle sostanze chimiche dei cibi sono concentrate soprattutto a livello del poro gustativo, che è il punto massimo dove convergono tutti gli stimoli che si devono legare alle molecole. Il meccanismo è tale per cui a livello di ciascun recettore ci saranno proteine recettrici che si collocano nella porzione apicale della cellula. Si distingue una parte basale e una apicale. È nella porzione apicale che si ha il legame della sostanza con le proteine recettrici. Il legame può avvenire o attraverso l'apertura di canali ionici o ci può essere l'innesco di reazioni metabotrope che hanno comunque come risultato finale la depolarizzazione della cellula, che rilascia neurotrasmettitore e insorgenza di potenziale di recettore. A differenza di quelli olfattivi non generano potenziale d'azione, ma di recettore. Anche nel caso delle cellule contenute nei bottoni gustativi, si tratta di cellule che si riproducono continuamente, addirittura ogni due settimane. Le cellule ancora non differenziate che si adranno a sostituire con i recettori si trovano già a livello dei bottoni. I circuiti principali sono l'ipotalamo e l'amigdala, utili per le sensazioni di sazietà e appetito e aspetti affettivi come gradevole/sgradevole. Le afferenze che arrivano dai diversi nervi cranici convogliano sul bulbo e poi sui talamo e da qui insula e corteccia frontale, ipotalamo e amigdala che sono strettamente connessi col nucleo del tratto solitario. Quando si parla di corteccia gustativa si fa riferimento a una porzione frontale, sottoregione ristretta, che elabora essenzialmente informazioni gustative. Esiste il fenomeno della fame specifica, esaminato in laboratorio o nell'esperienza quotidiana, quando si ha una fame per qualcosa di specifico. Si ha quando si tengono gli animali con una dieta deprivata per uno specifico alimento nutriente. Dopodichè si lasciano gli animali liberi di scegliere e mangiare diversi alimenti presentati. Si vede come essi scelgono l'alimento che preferiscono. Questo indica che l'elaborazione del gusto influenza la nostra assunzione di cibo e acqua regolando i sensi di sazietà. Un'altra cosa scoperta è stata che sottoponendo gli animali a esperimenti di condizionamento. Si presente all'animale un gusto e un suono provenire dalla stanza e subito dopo si ha la presentazione di una sostanza nauseante. Questo primo esperimento ha registrato che questi animali, dopo una serie di training di questo tipo gusto-sostanza nauseante-tono, essi avevano un'avversione per il gusto ma nessuna associazione con il tono. Quando la sostanza nauesante è stata sostituita da una scossa elettrica si è visto che si aveva un'associazione con il tono e nessuna associazione con il gusto. Questo indica che l'avversione appresa per gli stimoli gustitativi (ma non altri tipi di stimolo) sono associati ad esperienze sgradevoli (nausea). Non c'è invece condizionamento gustativo in associazione con esperienze di dolore/paura. 18. Sistema motorio Il sistema motorio, oltre a regolare i movimenti del nostro corpo, ci permette di tenere una certa postura. C'è inoltre una parte di sistema motorio detto autonomo che permette di controllare i movimenti che avvengono grazie alla muscolatura liscia all'interno del nostro organismo. Una definizione di sistema motorio è quella di un insieme di strutture cerebrale e vie nervose che controllano contrazione muscolare, al fine di regolare la postura (posizione del corpo nello spazio) ed eseguire movimenti (volontari, riflessi, ritmici). I neuroni fanno sinapsi con i muscoli, quindi anche i muscoli sono cellule eccitabili che grazie all'attivazione si contraggono e permettono il movimento. Le aree cerebrali, in linea generale, che fanno parte del sistema motorio, generano movimento attraverso le connessioni col midollo spinale, in quanto qui c'è la via d'uscita dei codici neurali che andranno a stimolari i muscoli. Questo avviene grazie ai corpi cellulari dei neuroni (motoneuroni) che si trovano nelle corna ventrali della sostanza grigia del midollo spinale. Con il loro assone attivano i muscoli. L'assone esce, passando attraverso la radice ventrale, che si unirà alle fibre che passano attraverso la radice dorsale in quello che viene chiamato nervo spinale. I nervi spinali sono allora sistema nervoso periferico nel caso delle efferenze. Una fibra muscolare lunga va da un tendine, che permette al muscolo di essere attaccatto all'osso, a un altro, riceve la sinapsi da un solo motoneurone. Un motoneurone fa sinapsi con più fibre muscolari, perchè l'assone forma ramificazioni. Si tratta di una sinapsi che in questo caso prende il nome di giunzione terminale, attraverso rilascio di neurotrasmettitore (acetilcolina). L'acetilcolina è rilasciata quando il motoneurone scarica una serie di potenziali d'azione, che arrivano a livello del terminale sinaptico (sinapsi chimica). Le vescicole si fondano con la membrana sinaptica e il neurotrasmettitore viene rilasciato. I suoi recettori si legano con esso determinando un potenziale d'azione anche nella fibre del muscolo, il quale le fa contrarre. La conseguenza del rilascio del neurotrasmettitore è quindi contrazione delle fibre. Si definisce unità motoria l'insieme del motoneurone e delle fibre muscolari da esso contattate e attivatate. Parliamo di unità motoria perchè i muscoli possono avere un numero diverso di unità motorie e questo può avere funzione diverse. Questo ci permette di graduare più o meno la forza che serve per contrarre un muscolo. Ci sono muscoli la cui contrazione può essere graduata in maniera fine e lieve (come i muscoli della dita, quelli extraoculari), altri in cui ciò non avviene e questo dipende dalle unità motorie. Per muscoli che compiono movimento grossolani (come la coscia) si avranno un numero di unità motorie minore, cioè un motoneurone innerva tante fibre. Per esempio: muscoli dalla gamba 1:2000. Muscoli extraoculari: 1:10. Quando i motoneuroni sono attivi i muscoli si contraggono. Se il muscolo considerato è il bicipite, il braccio tira verso l'alto avambraccio, flettendosi. Supponiamo ci sia una leva: quando il muscolo si contrae tira verso di sè l'osso, facendo piegare l'articolazione. I muscoli quindi sono legati alle ossa grazie ai tendini. L'estensione si ha quando il muscolo del tricipite (detto antagonista) si contrae. Quando questo si contrae, tira l'avambraccio, che si distende. Flessione ed estensione avvengono a livello di tutte le articolazioni. Quando ciò avviene si ha il movimento. Quando un'agonista si contrae, l'antagonista dev'essere rilassato. I muscoli sinergici contribuiscono a generare lo stesso movimento. L'effetto di una contrazione muscolare non è sempre l'accorciamento di un muscolo (come nel caso del braccio). Distinguiamo tra contrazione isotonica, in cui il muscolo si accorcia e contrazione isometrica, in cui il muscolo non cambia di lunghezza. Isotonica, dal greco, forza costante e isometrica, lunghezza costante. Il motoneurone che scaricava a una certa frequenza, quando si ha contrazione isotonica, viene fatta aumentare la frequenza di scarica, simboleggiata da barre più vicine tra di loro. Nella fase di mantenimento del piegamento avvenuto la frequenza di scarica diminuisce. C'è dunque una scarica fasica e una scarica tonica, cioè costante, per far mantenere la posizione. In questo caso cambia la posizione (il muscolo si accorcia) ma la forza esercitata è costante durante l'accorciamento. La contrazione isometrica rimane costante la lunghezza del muscolo. Questo è possibile quando viene applicato un peso. Supponiamo di applicare un peso su una mano. Il braccio non si piega, ma il muscolo si contrae senza accorciarsi. La forza non rimane costante, ma aumenta. I motoneuroni scarica in maniera simile alle contrazioni isotoniche, nonostante l'effetto finale sia diversi. Partendo dalle aree motorie della corteccia cerebrale (area motoria primaria, area premotoria e area supplementare motoria). Abbiamo una corteccia motoria destra che permette la contrazione dei muscoli a sinistra e viceversa. Si passa poi al tronco dell'encefalo, al midollo spinale e generazione del movimento. Questa è la via più semplice, via in cui si nota un'organizzazione gerarchica, cioè queste strutture sono parte di una gerarchia. I punti alti della gerarchia si occupano di pianificare la strategia e i punti bassi si occupano di eseguire il movimento. Nella corteccia cerebrale abbiamo gli aspetti strategini del movimento: programma motorio (movimenti volontari), ma anche regolazione di stazioni inferiori. Il tronco encefalico è importante per l'integrazione degli stimolo visivi, vestibolari e somatosensoriali. Si occupa del controllo della postura. La stazione inferiore del midollo spinale, con i suoi motoneuroni, rappresenta la via d'uscita in cui tutti i comandi motori vi convergono. Il midollo spinale ha anche un controllo motorio autonomo perchè ci permette di generare riflessi. C'è anche un'organizzazione in parallelo perchè non esiste una sola via ma molteplici, che controllano diversi aspetti del movimento. Infatti la corteccia cerebrale può essere connessa al tronco dell'encefalo, ma ce n'è un'altra che la collega direttamente al midollo spinale. Altre strutture del sistema motorio non collegate direttamente come via efferente al midollo spinale non determinano direttamente un movimento perchè non sono connesse al midollo spinale, ma sono comunque importanti perchè collaborano alla pianificazione e generazione dei movimenti. Tra queste abbiamo i gangli della base, nell'encefalo, sotto la corteccia. Questi inviano informazioni alla corteccia motoria e ricevono informazioni dalle aree corticali. Il cervelletto invece serve a farci eseguire i movimenti in maniera coordinata e a correggere il movimento se viene eseguito in maniera sbagliata. È inoltre sede della memoria e dell'apprendimento. Le sue afferente sono ricevute dalla corteccia e le manda al tronco cerebrale. Può riceverle anche dal midollo spinale. Quando si impara un nuovo movimento, come uno sport o la bicicletta, non si è stabili perchè compiamo movimenti sbagliati, che vengono corretti con la pratica. Di questo se ne occupa il cervelletto. Le informazioni sensoriali, grazie ai recettori sensoriali, sono importanti perchè senza di essi non potremmo pianificare ed eseguire correttamente nessun movimento. Nel caso di una malattie muoiono i neuroni dei gangli delle radici dorsali; questi pazienti non ricevono più informazioni sensoriali, per esempio potrebbero non percepire di avere qualcosa in mano. Quindi informani sullo spazio esterno, posizione del corpo, stato dei muscoli (visive, uditive, somatosensoriali, propriocettive e vestibolari) sono essenziali per l'esercuzione del movimento. Ciò avviene tramite un controllo a retroazione (a feedback) o anticipatorio (feedforward). Nel primo caso le informazioni sensoriali arrivano a un centro comparatore, come il cervelletto, e ci dicono come sta avvenendo il movimento. Il centro comparato paragona come sta avvendo il movimento a come dovrebbe essere (piano d'azione). Questo avviene, per esempio, mentre si cammina su una barca: il movimento non è stabile e dobbiamo continuamente modificare il nostro movimento per rimanere in equilibrio. Il controllo a retroazione avviene grazie ai livelli bassi della gerarchia come cervelletto, tronco, midollo spinale. Se il movimento è ballistico, cioè veloce, non riusciamo a correggerlo nonostante ci sia l'informazione sensoriale. Per esempio, quando voglio colpire una mosca con la mano e questa scappa, non faccio in tempo a correggere il movimento per il mio scopo. Il controllo anticipatorio del movimento da parte delle sensazioni ci permette di programmare il movimento nella maniera più adeguata, come il movimenti di un giocatore di baseball che deve ricevere una palla e si prepara adeguatamente al movimento. Questo controllo avviene grazie ai livelli alti della gerarchia, come corteccia e gangli della base. Diamo la definizione di due tipi di muscoli che ci permettono di eseguire movimento in quanto tutte le strutture del sistema motorio si dividono in due parti a seconda del movimento che determinano. Abbiamo muscoli che definiamo distali (più lontani dall'asse corporeo), come i muscoli delle dita, della mano o del braccio, che ci permettono movimenti fini e precisi rispetto ai muscoli che definiamo assiali (più vicini all'asse corporeo) o prossimali, deputati al controllo dell'equilibrio e della postura. Ci sono strutture del sistema motorio che fanno parte dei sistemi mediali (che si occupano delle contrazioni di muscoli distali) o dei sistemi laterali (che si occupano delle contrazione di muscoli assiali). Nelle corna di sinistra della parte ventrale del midollo spinale ci sono motoneuroni in posizione più esterna alla linea mediana che controllano i muscoli distali degli altri e sono responsabili dei movimenti volontari; fanno parte dei sistemi laterali. Nella parte sinistra del midollo spinale ci sono altre due divisioni: nella parte ventrale ci sono neuroni che si occupano di muscoli flessori e nella parte dorsale di muscoli estensori. Non ci sono solo motoneuroni nelle corna del midollo spinale. Ci sono anche interneuroni il cui assone non esce dalla regione in cui si trovano. Sia motoneuroni che interneuroni ricevano informazioni sensoriali e informazioni dalla corteccia. Gli interneuroni fanno parte di sistemi mediali e laterali. Si trovano in una delle due zone viste a livello del midollo spinale. Gli interneuroni in questo caso si chiamano propriospinali: questi connettono più segmenti. Si distinguono in due categorie: quelli dei sistemi laterali contribuiscono al controllo di pochi muscoli alla volta, mentre quelli dei sistemi mediali contribuiscono al controllo di parecchi muscoli in maniera coordinata (postura). Perchè si abbia una postura eretta è necessario infatti che si abbiamo sempre contatti più muscoli nello stesso momento e in entrambi i lati della linea mediana. Questo avviene grazie all'attivazione di più segmenti del midollo spinale grazie agli interneuroni che li connettono. Per i movimenti fini sono necessarie poche zone e il movimento è unilaterale. Anche nel tronco encefalico abbiamo sistemi laterali e mediali. Abbiamo gruppi di neuroni che fanno parte o di uno o dell'altro sistema, cioè che si occupano o della contrazione dei muscoli distali o prossimali. I nuclei presenti nel tronco cerebrale danno origine a dei tratti di assoni. Il tratto rubrospinale fa parte dei sistemi laterali; questo deriva dal nucleo rosso. Questo serve a farci controllare i muscoli delle braccia. Nell'uomo questo tratto è molto poco sviluppato rispetto ad altri animali. Dei sistemi mediali fanno parte i tratti: reticolo-spinale, vestibolo-spinale, tetto-spinale. Il primo deriva da sostanza reticolare e arriva a midollo spinale. La sostanza reticolare è un insieme di neuroni che si trovano nel tronco cerebrale che servono al controllo della postura e infatti fa parte dei sistemi mediali. Il tratto vestibolo-spinale deriva da nuclei vestibolari nel tronco centrale che fanno sinapsi nel midollo spinale. Il tratto tetto-spinale deriva dal mesencefalo (da una zona che si chiama tetto); serve per il controllo di movimenti che ci permettono di seguire lo stimolo che si muove, muovendo per esempio testa e occhi, in modo che lo stimolo cada sempre nella fovea. Si dice che il reticolo-spinale e il vestibolo-spinale sono diretti, cioè non crociano, mentre il tetto-spinale è crociato, passando la linea mediale. Ci sono poi sistemi a proiezione diffusa, insieme di neuroni nel tronco cerebrale; hanno assoni e contatti sinaptici diffusi. Servono a modulare e regolare l'eccitabilità di altri neuroni. In ordine gerarchico, c'è la corteccia cerebrale, che ha anch'essa tratti che appartengono all'uno o all'altro sistema. La differenza in questo caso è che la corteccia si occupa di movimenti volontari. L'area motoria primaria e l'area premotira danno origine al tratto cortico-spinale, che fanno sinapsi dalla corteccia al midollo spinale. Ci sono tratti cortico-spinale laterale, che attivano motoneuroni del midollo nei sistemi spinali leterali. Il tratto cortico-spinale crocia, cioè passa dall'altra parte della linea mediale. Il tratto cortico-spinale ventrale invece fa parte del sistema mediale: esso attiva i motoneuroni del midollo spinale nei sistemi mediali. Questo tratto è diretto, ma resta ipsilaterale. Esiste anche il tratto cortico-bulbo-spinale, cioè la corteccia ha neuroni con assoni che fanno sinapsi nel tronco. 19. Riflessi spinali Ci sono tre tipi di movimento: volontari, riflessi e ritmici. I primi sono quelli che dipendono dalla nostra volontà, sono sempre finalizzati, per la maggior parte appresi; la loro precisione aumenta con l'esercizio e, una volta appresi, non richiedono partecipazione cosciente. I movimenti riflessi sono involontari, rapidi, stereotipati, innati e modulati da stimolo. Un tipico movimento riflesso è quello che avviene battendo il martelletto nel ginocchio. I movimenti ritmici, come la masticazione o la deambulazione sono un insieme di movimenti volontari e riflessi, spesso innati ma hanno inizio e fine volontari. I circuiti che ci permettono di generare riflessi spinali sono nel midollo spinale. Se c'è uno stimolo, che viene recepito da un recettore, esso manda l'informazione al midollo spinale grazie ad afferenze che sono assoni di neuroni il cui corpo cellulare è il ganglio delle radici dorsali. Nella generazione del riflesso il motoneurone fa contrarre un muscolo chiamato effettore che determina la risposta motoria. I riflessi sono o monosinaptici (c'è una sola sinapsi tra neurone sensoriale e neurone motorio) o polisinaptici (sono costituiti da più sinapsi presenti nel centro del riflesso, nel midollo spinale). I propriocettori sono recettori all'interno dei muscoli che segnalano la lunghezza o la tensione. La capacità di acquisire informazioni che riguardano lo stato dei muscoli e quindi su come il nostro corpo è posto e si muove nello spazio è detta propriocezione. I propriocettori sono anche responsabile della generazione di certi riflessi. I propriocettori hanno dunque due funzioni principali: fornire informazioni sensoriali sullo stato dei muscoli e proteggere i muscoli, grazie ai riflessi. Abbiamo due tipi di recettori per propriocezione: fuso neuromuscolare, che si trovano tra le fibre muscolari; si dice che sono in parallelo, cioè si trovano disposte parallelamente alle fibre muscolari che servono alla contrazione. Il fuso neuromuscolare serve a recepire la lunghezza del muscolo. Tra muscolo e tendine c'è l'altro tipo di propriocettore: organi tendinei del Golgi. Essi sono situati in serie e ci danno informazioni sullo stato di tensione dei muscoli. I fusi sono contenuti in vere e proprie capsule; sono fibre muscolari modificate attorno alle quali sono avvolti gli assoni sensoriali. Quelle nel fuso vengono chiamate fibre muscolari intrafusali e quelli e che si trovano al di fuori sono chiamate extrafusali. Quando il muscolo si allunga, il fuso scarica potenziale d'azione a maggiore frequenza, mandando un segnale che indica che il muscolo si è stirato. Questo avviene perché gli assoni sensoriali avvolti alle fibre muscolari intrafusali vengono stirate anch'esse col muscolo: meccanicamente la membrana plasmatica degli assoni si tira; sulla membrana i canali per il sodio si aprono, entra più sodio e si genera potenziale d'azione. In questo modo il fuso neuromuscolare dà informazioni sulla posizione delle varie parti del corpo, sul loro movimento e velocità. Oltre agli assoni muscolari avvolti c'è un altro tipo di innervazione che riguarda i due estremi del fuso, che ricevono fibre motorie. I motoneuroni che fanno sinapsi sul fuso (chiamati gamma) servono a regolarne la lunghezza del fuso e quindi la sua sensibilità. Le fibre muscolari del fuso che si trova nel muscolo è molle in quanto il muscolo si contrae e scarica potenziale d'azione. Le fibre dei motoneuroni gamma fanno contrarre le parti estreme del fuso, così la parte centrale si tende. Il fuso è così pronto a rispondere a un minimo stiramento del muscolo. Grazie a entrambi i motoneuroni, alfa e gamma, il fuso rimane teso nonostante il muscolo si accorcia. In questo modo anche se il muscolo è contratto il fuso è sensibile in maniera fine allo stiramento. Quando lo stiramento è eccessivo o improvviso e si potrebbe strappare, si genere un riflesso miotatico per far contrarre il muscolo in modo che non si danneggia. È generato dal fuso neuromuscolari che manda il segnale di stiramento. Le fibre all'interno del fuso sono le fibre muscolari. Quando il muscolo si allunga ed è eccessivo il fuso neuromuscolare lo segnala con una scarica. C'è una sinapsi del neurone sensitivo primario sul motoneurone, che si attiva e scarica a maggiore frequenza. Questo ne determina la contrazione, cioè si accorcia. Questo tipo di processo molto rapido è monosinaptico. Per esempio: abbiamo un bicchiere in mano e viene versato dentro qualcosa. Generalmente il braccio dovrebbe distendersi, ma per riflesso il braccio si contrae. Si tratta di uno stiramento passivo. Un altro esempio di riflesso miotatico è quello della risposto al colpo del martelletto a livello della rotula che determina l'estensione della gamba. Quando il martelletto colpisce il legamento, questo colpo crea una forza che tira il muscolo stirandolo. Perchè un riflesso avvenga in maniera fluida ed efficace non basta la contrazione dei muscoli stirati eccessivamente; l'ideale è l'inibizione dei muscoli antagonisti. Questo avviene grazie allo stesso circuito del riflesso in cui la fibra sensoriale manda l'informazione si stiramento anche ad un interneurone inibitorio, che inibisce il motoneurone alfa che fa sinapsi sul muscolo antagonista. Questo processo si chiama inibizione reciproca. Il riflesso miotatico dunque: consente di contrarre muscoli se stirati troppo o in maniera improvvisa (riflesso di stiramento); di correggere errori durante l'esecuzione di movimenti; di mantenere tono muscolare (tensione dei muscoli in stato di risposo e forza con cui i muscoli si oppongono a stiramento); opporsi alla forza di gravità e mantenere la postura. I riflessi spinali avvengono grazie a circuti nel midollo spinale, ma c'è un continuo controllo da parte dei centri superiori, di tipo discendente, che avviene da corteccia e tronco. Questo controllo permette di regolare sensibilità. A seconda delle situazioni abbiamo un tono muscolare e dei riflessi diversi. Se siamo in piedi rilassati o camminiamo avremo un certo tipo di tono muscolare e di estensione del fuso. Camminando all'interno di un autobus in movimento in realtà teniamo i fusi neuromuscolari più pronti a rispondere. Si dice che il riflesso può essere facilitato o inibito a seconda del fatto che siamo in situazioni di relax o no. Questa relazione di riflesso avviene grazie ai centri superiori, in quanto c'è un programma motorio che va a dire ai motoneuroni gamma di regolare la lunghezza dei fusi a seconda delle situazioni. Se durante un movimento c'è un errore o una perturbazione, si genera uno stiramento, i fusi scaricano e generano un movimento riflesso. Se i motoneuroni gamma sono molto attivi, al minimo stiraramento c'è una grossa contrazione; c'è un cosiddetto guadagno del riflesso, che in questo caso è elevato, cioè c'è un reclutamento di un numero elevato di motoneuroni alfa. L'organo tendineo del Golgi, posizionato in serie, si trova nel tendine. Nel tendine ci sono le fibre collagene e intrecciate a queste ci sono le ramificazioni di un assone sensoriale. Questa serve a segnalare quando c'è una tensione del muscolo attaccato al tendine. Quando il muscolo si accorcia (e cioè c'è maggiore tensione muscolare) l'organo tendineo del Golgi scarica. C'è maggiore tensione nella contrazione isometrica. L'aumento di tensione da sì che i tendini vengono tirati e le membrane di queste fibre presentano canali per il sodio che meccanicamente si aprono; così il sodio aumenta generando potenziale d'azione. C'è un riflesso quando il muscolo si contrae eccessivamente. Il riflesso sarà di disattivazione della contazione eccessiva o prolungata: il muscolo si rilassa evitando danneggiamento. Il circuito in questo caso è polisinaptico e il riflesso miotatico è di tipo inverso. Il riflesso flessorio è un riflesso di tipo spinale è quello che ci fa ritrarre (flettere) un muscolo a contatto con qualcosa di doloroso. Questo tipo di riflesso ci permette di allontanare uno stimolo dolorifico, con funzione di protezione. I nocicettori fanno sinapsi con più interneuroni, che si attivano; alcuni sono eccitatori e altri inibitori. Un interneurone eccitatorio fa contrarre i muscoli che determinano la flessione; un interneurone inibitorio inibisce i muscoli che determirebbero un'estensione. Ci sono interneuroni che crociano, cioè una volta ricevuta un'informazione a destra la portano a sinistra. Abbiamo dunque flessione ipsilaterale ed estensione controlaterale (in modo che un arto si flette, in modo da allontanarsi dalla fonte di dolore, ma l'altro si deve estendere in modo da mantenere la postura e l'equilibrio). I centri superiori modulano i riflessi grazie a delle efferenze verso i centri del riflesso. Se c'è una disconnessione parziale o totale dei centri dei riflessi alle regioni superiori e le fibre non arrivano più. Le fibre sono disconnesse, quindi si ha una perdita definitiva della sensibilità e del movimento volontario. Se c'è una disconnessione, i riflessi sono inizialmente aboliti (shock spinale) e compaiono riflessi che non avevamo (riflesso di Babinsky). I movimenti ritmici hanno una certa ripezione, come la deambulazione o la masticazione. Nella deambulazione una gamba si flette e l'altra si estende, quindi è un po' simile al riflesso flessorio, anche se non è determinato da un dolore. Le vie discendenti danno il comando di inizio e fine di una serie di circuti secondo cui alcuni muscoli si flettono e altri si estendono. Tutto ciò è dovuto ai circuiti locali spinali o meglio generatori centrali di schemi motori, cioè neuroni che indipendentemente dalla nostra volontà generano scariche che ci fanno contrarre certi muscoli e flettere altri e ci permettono di camminare. Questo gruppo di neuroni si trovano nel midollo spinale. Questo è stato capito e studiato dai gatti, in cui è stata operata una disconnessione nel midollo spinale dai centri superiori. Egli è in grado di camminare e questo dimostra che i movimenti ritmici non dipendono dalla corteccia; è compromessa però la volontà, cioè il gatto non è in grado di iniziare a camminare, ma è in grado di farlo ponendolo su un tapis-roulant, per esempio. I generatori centrali di schemi motori si trovano uno per arto e posso anche operare indipendentemente dagli altri arti, indipendenti dalle vie discendenti e quindi dalla voltà; sono dunque involontari e innati. Le vie discendenti invece regolano inizio e fine dei movimenti ed equilibrio. La postura è la posizione che assume il corpo a riposo o in movimento e delle sue parti. I circuiti corticali ci permettono di fare un programma motorio e tra questi programmi ci sono anche quelli per gli aggiustamenti posturali, che inviano il comando a regioni del tronco cerebrale. Questi comando sono poi eseguiti da centri del tronco encefalico, tra cui la sostanza reticolare, insieme di neuroni sparsi per il tronco cerebrale, che controllano i riflessi spinali e soprattutto il riflesso miotatico. In esperimenti sui gatti è stato visto che bloccando farmacologicamente la sostanza reticolare si perde la postura. Importante nel controllo della postura è l'insieme delle afferenze sensoriali, sia quando dobbiamo fare movimenti posturali, sia come quando, in risposta a un suono, il soggetto deve tirare una leva: non c'è solo il movimento volontario del bicipite, ma anche il movimento posturale che faccia in modo che il soggetto non cada. Il programma viene dalla corteccia motoria, che informa il midollo spinale per il movimento volontario, ma anche la sostanza reticolare, che esce sempre dal midollo spinale. Grazie al midollo spinale avremo sia la contrazione dei muscoli per il movimento volontario sia l'aggiustamento dei muscoli per il movimento posturale. Le informazioni sensoriali esercitano un controllo a feedback, cioè non solo sul movimento, ma anche sulla postura. Quando si cammina su una barca si fanno continuamente aggiustamenti posturali. Questo avviene integrato le informazioni sensoriali che formano un centro comparatore, che integrano le informazioni sensoriali con il programma motorio, che danno via al movimento a e agli aggiustamenti posturali e che può anche essere corretto nel caso in cui l'aggiustamento è errato. La postura è quindi regolata dalla sostanza reticolare, che è nel tronco cerebrale, con le sue connessioni al midollo spinale, che invece è rappresentato in sezioni. Tra i vari tratti propri dei sistemi mediali c'era il tratto reticolo-spinale, che si origina dalla sostanza reticolare e fa sinapsi nel midollo spinale. La sostanza reticolare regola la postura aumentando l'efficienza con cui i motoneuroni alfa e gamma del midollo spinale rispondono agli stimoli, pertanto aumenta l'efficienza del fuso neuromuscolare (informazioni sulla lunghezza dei muscoli, cioè sulla posizione delle varie parti del corpo) e aumenta l'efficienza del riflesso miotatico (tono muscolare e postura).
Scaricare