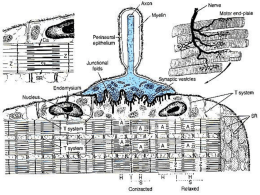
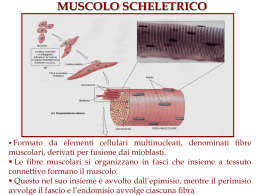
Muscolo Liscio Striato Scheletrico Slow twitch Cardiaco Tonico Fasico Fast twitch www.fisiokinesiterapia.biz Cellula comune Nucleo Membrana Citoplasma Mitocondri Cellula del muscolo striato= una fibra muscolare Multinucleato Sarcolemma Sarcoplasma Sarcosomi Muscolo scheletrico: la cellula è spesso lunga quanto il muscolo, con diametri da 10 a 100 micron (anche se un muscolo atrofizzato è più spesso) perimisium muscolo fascicolo endomisium fibra Un muscolo è fomato dalla fusione di centinaia di fibre. Gran parte dell’interno della fibra muscolare è formata da filamenti proteici , le cosiddette“miofibrille”. Muscolo Scheletrico: attaccato alle ossa tramite i tendini. Serve per produrre movimenti o esercitare forze. Serve per locomozione, postura, respirazione, mangaire, espressione…. I muscoli sono innervati da motoneuroni, comandati dal sistema nervoso centrale. L’unita’ motoria comprende il neurone, e le fibre muscolari che lui attiva. La risposta graduata è dovuta alla variazione nel grado di attivazione del tessuto dai nervi. Il muscolo si chiama striato perchè visto sotto microscopio, forma delle bande chiare e scure alternativamente ripetute. Le bande sono allineate in senso transverso. Miofibrilla: lunga quanto la cellula, e allineata con altre miofibrille. Diametro, circa 2 micron L’unita contrattile: il sarcomero Z A Z I H I Durante la contrazione: la distanza tra le linee Z diminuisce, I diventa più piccolo, H sparisce, ma A rimane costante. A: Scura, anisotropa I: chiara, isotropa Z: disco Z, o linea H: nel centro della zona A, è chiara Lunghezza: 2.5 micron Durante la contrazione: la distanza tra le linee Z diminuisce, I diventa più piccolo, H sparisce, ma A rimane pressochè costante Z A I H Z A Z Z Il sarcolemma, la membrana esterna che circonda ogni fibra, ha il ruolo di permettere la propagazione dell’onda di potenziale su tutta la superficie della cellula Organizzazi one molto ordinata Le miofibrille contengono due tipi di filamenti, formati da due tipi di proteine. Filamento spesso di diametro 10-12 nm, di miosina. Filamento sottile di diametro 6-7 nm, di actina. I filamenti spessi hanno delle proiezioni ogni 40 nm. Secondo la teoria degli “sliding filaments ” o scorrimento dei filamenti, questi si sovrappongono e le proiezioni formano i cosiddetti “cross-bridges” Per ricordare: Muscolo Fascicolo Fibra muscolare Miofibrilla Miofilamenti Filamenti sottili: soprattutto actina Filamenti sottili: soprattutto miosina Le cellule sono multinucleate perché sono formate dalla fusione di cellule precursori. Non possono dividersi. I muscoli possono ipertofizzarsi, ma questo è causato da un aumento del diametro delle fibre. Filamenti sottili: Contengono le proteine actina, troponina e tropomiosina. La proteina F-actina è composta da due catene intrecciate ad elica. Una molecola di ADP è attaccata ad ogni catena. La tropomiosiona è infilata dentro l’F-actina e ha tre subunità: una che si lega all’actina, una alla tropomisiona, e una che si lega in maniera reversibile con lo ione Ca++. Filamenti Spessi: la miosina è composta da due catene pesanti (meromiosina pesante) e 4 leggere (meromiosina leggera). Ogni filamento spesso è composto da centinaia di molecole di miosina. La miosina leggera si aggrega per formare filamenti, mentre la miosina pesante, che è la parte attiva, forma due teste piegate. Le teste si legano con l’actina e funzionano da ATPasi (idrolizzano ATP in ADP +P rilasciando energia). L’energia viene utilizzata nel meccanismo di contrazione. La contrazione muscolare richiede tanta energia, e il muscolo scheletrico contiene tanto ATP, che viene prodotto dai mitocondri durante i periodi di riposo. Nel sarcoplasma vi sono anche abbondanti riserve lipidiche e di glicogeno. Se le riserve di ATP finiscono durante I periodi di contrazione prolungata, l’ADP generata dall’idrolisi dell’ATP viene riconvertito in ATP attraverso glicolisi anerobica, che porta all’accumulo di acido lactico. Le fibre muscolari sono circondate da una rete di elementi vescicolari: il reticolo sarcoplasmatico che regola i meccanismi di contrazione attraverso il rilascio (che porta alla contrazione) e il richiamo (che porta al rilassamento) di Calcio. Lo ione Ca++ viene rilasciato quando c’è una depolarizzazione che apre i canali ionici del reticolo sarcoplasmatico. La teoria dei filamenti scorrevoli (Huxley 1954) La teoria modellizza la contrazione come dovuta al movimento relativo tra i filamenti spessi e i filamenti sottili . L’osservazione è basata sul fatto che le bande A rimangono invariate durante una contrazione. Lo scorrimento è dovuto alla formazione e alla rottura di ponti tra actina e miosina. Le proiezioni sulle fibre di miosina si attaccano al sito attivo di actina e lo tirano verso l’interno del sarcomero. Dopodiché la miosina lo rilascia e si attacca al prossimo sito per ripetere il ciclo. Durante il riposo, le interazioni tra actina e miosina sono inibite a livello del sito attivo di interazione dei due filamenti: è ostruito dal complesso troponina-tropomiosina. Allorché un potenziale di azione si propaga sulla fibra, il reticolo sarcoplasmatico rilascia il Ca++. Il Ca++ si lega alla troponina, attivando un cambiamento conformazionale della molecola, che a sua volta espone il sito attivo dell’actina, al quale si lega la miosina. Prima che inizi la contrazione, l’ATP si lega alla testa della miosina. Questo viene subito idrolizzato dall’ATPase sulla testa. ADP e P rimangono attacate alla testa. A questo punto, grazie all’energia fornita dall’ATP e con il rilascio di P, la testa di miosina si ruota (questo è il power stroke), causando uno scorrimento del filamento di actina. Una volta che la testa si ruota (o comunque varia l’angolo), l’ADP viene rilasciato, e un nuovo ATP può’ legarsi alla testa. Senza ATP, non c’è rilascio. Quindi l’ADP è necessario affinché si leghino, e ATP perché si separino. Se la teoria è giusta, si dovrebbe osservare un movimento di miosina durante una contrazione. Dai dati più’ recenti con diffrazione a raggi X, sembra che non ci sia una rotazione, ma un effettivo accorciamento dei filamenti spessi (la teoria invece è basata sul fatto che i filamenti non si accorciano ma si muovono uno rispetto all’altro, quindi le bande A rimangono invariate). Adesso la versione generalmente accettata punta su una contrazione delle catene leggere di miosina, ma sempre con lo scorrimento dell’actina. Il problema con questa teoria (che è degli anni 50) è che ogni volta che si trova un’inconsistenza, viene proposto un meccanismo alternativo, pur basandosi sempre sul concetto dei filamenti scorrevoli. Una teoria alternativa proposta da Pollack et altri afferma che i filamenti di miosina sono interconessi tra di loro, e l’actina scorre al loro interno, agganciandosi ai siti di miosina una alla volta. Questo è accompagnato da un accorciamento della catena leggera della miosina. La base di questa teoria è che tutto dipende dalla struttura dell’acqua nel sistema. L’ingresso di Ca++ causa l’espulsione di acqua, come anche l’ATP. Questo è la causa principale della contrazione. Nello studio delle proprietà contrattili abbiamo le variabili: tempo, t lunghezza normalizzata rispetto a quella di riposo, (L/Lo) forza per unità di area, F velocità, v=dL/dt stiffness, dF/dl compliance, dL/dF lavoro, W=Fdl potenza, p=dW/dt I modi di definire una contrazione sono i seguenti: Isometrica: lunghezza fissa, la tensione aumenta con l’attivazione Isotonica: il carico è fisso, e il muscolo può accorciarsi Isocinetica: la velocità di contrazione è fissa, e la tensione può variare Contrazione isometrica: Il muscolo viene mantenuto a lunghezza costante, e sviluppa una forza quando stimolato elettricamente. Questo produce una contrazione o twitch. La forza varia con la lunghezza iniziale. Infatti, la tensione massima viene sviluppata quando il muscolo è a riposo. Il massimo si ha quando i due filamenti sono sovrpaposti per tutta la loro lunghezza, e quindi il numero di ponti è massimo. Twitch: il potenziale di azione dura 1-2 ms lo La tensione massima si ha quando i due filamenti sono sovrapposti per tutta la loro lunghezza, e quindi il numero di ponti è massimo. Se un muscolo viene stimolato, e un secondo stimolo viene applicato prima che il rilassamento venga completato, c’è una seconda contrazione, che sviluppa una tensione maggiore. Questo può essere dovuto al Ca++ che rimane dalla prima contrazione che va a sommarsi con la seconda onda di Ca++. Se lo stimolo viene ripetuto a una frequenza elevata il muscolo non riesce a rilassarsi, ma rimane in uno stato di contrazione. Questo stato si chiama tetano muscolare. velocità dello stimolo 100/sec Forza di contrazione 60/sec 40/sec 25/sec 15/sec 10/sec millisecondi Rapporto Lunghezza-Tensione La tensione sviluppata in un muscolo dipende dalla sua lunghezza. Possiamo evidenziare due tipi di tensione: •Tensione passiva, dovuto all’allungamento •Tensione attiva, sviluppata dal interazione actina-miosina. La tensione totale è la somma delle due. Qui viene applicato uno stimolo tetanico, s, per diversi allungamenti, ΔL La tensione passiva è dovuto alle proprietà meccaniche del tessuto connettivo. totale Un muscolo allungato è più’ sensibile al Ca++. Al contrario, uno accorciato lo è di meno (cioè ci vuole più’ alta concentrazione di Ca++ per farlo contrarre ancora). Contrazione isotonica Quando il muscolo tira un peso, la tensione inizia a aumentare perché si contrae prima in maniera isometrica. Quando la tensione è uguale al carico, il muscolo si contrae in maniera isotonica. La velocità di accorciamento dipende dal carico. mg Il carico ottimale è circa 1/3 della forza massima. La velocità massima di accorciamento è correlata con l’attivita’ dell’ATPase. Fibre veloci hanno ATPase più attivo, con alte velocità di regenerazione (o riciclo). Quando il carico è elevato, la velocità diminuisce perché il carico si oppone alla contrazione. Prima il muscolo deve utilizzare l’energia per superare la forza del carico, e poi usa l’energia rimasta per accorciarsi. Quando il carico è molto elevato, il muscolo non si può’ contrarre anche quando attivato. Se il carico supera la tensione isometrica massima , la fibra si allunga. mg Il rapporto forza- velocità è determinato da: • numero di “ponti” tra actina miosina (sovrapposizione) • quantita’ di Ca++ rilasciato dal reticolo sarcoplasmatico • attivita’ dell’ATPase • numero di fibre reclutate Una fibra ipertrofizzata ha effetivamente più sarcomeri in paralleo Aumenta la forza generata ma non cambia la velocità di accorciamento, né l’accorciamento massimo. Una fibra allungata (con lo streching!) ha più’ sarcomeri in serie non cambia la forza generata, ma aumenta la velocità e la capacita di contrarre. Fibre lenti Fibre veloci Fibre intermedie Produzione ATP Fosforilazione ossidativa (aerobico) Glicolisi (anerobico) Fosforilazione ossidativa (aerobico) Colore (mioglobina) mitocondri Intenso Tanti Chiaro pochi Medio Diametro fibra Piccolo grande Medio velocità di afatticamento Lento veloce medio Caratteristiche Mantengono attività tonica per lunghi periodi Piccoli istanti di attivita potente intermedie Muscoli agonistiantagonisti Muscolo Cardiaco E’ altamente vascolarizzato: 1 capillare per ogni cellula. Ha la proprietà di contrarsi spontaneamente, in maniera autoritimica. Il diametro delle miofibrille è di circa 15 micron, la lunghezza è di circa 85. Si hanno uno o due nuclei centrali. Ipertorfia muscolare: le cellule si gonfiano e allungano. Muscolo cardiaco dannegiato: il tessuto non si rigenera. Tanta Muscolo Liscio Si trova nei vasi, pelle e viscere. E’ un tessuto involontario, inoltre la cellula può contrarsi solo in parte. L’idrolisi di ATP è più lenta: le contrazione sono lente, prolungate e più efficienti (meno ATP richiesto). Ci sono due tipi di cellule SMC (Smooth Muscle Cells) : • Unità singole (viscere), che muovono in sincronia. L’attività è miogenica: cioè spontanea. In ogni caso sono innervate dal ANS (Sistema Nervoso Vegetativo ) • Unita multiple: non hanno attività spontanea e sono attivate da impulsi nervose dal ANS Le SMC mantengo uno stato di tensione costante, che può essere modulato da ormoni, fattori locali o attività autonoma. Le cellule si allungano e si accorciano molto di più delle striate. Ad esempio, la vescica, quando è piena ha una circonferenza di 30 cm. A vuoto, di 8 cm. Se era fatta da cellule striate, quanto può cambiare di dimensioni? scheletrico Cardiaco Liscio Diametro μm 50-100 5-10 2-10 Lunghezza μm centimetri 100-200 20-600 Nucleo tanti 1-2 1 Mitocondri 2-12% volume 40% volume 5% volume Tuboli diametro 20 nm 100 nm Non ci sono Reticolo sarcoplasmatico Abbondante e ben organizzato (9%) Spesso e sottile Poco (1-2%) <2% Spesso e sottile Spesso sottile e intermedie. Actina manca di troponina Innervazione CNS- voluntario Tutto o niente ANSinvoluntario ANS- involuntario Sarcomeri si si no Calcio Dal reticolo sarcoplasmatico Extra cellulare Caveolae (vescicoli sotto il sarcolemma) Calcio si lega a Troponina Troponina Calmodulina Rigenerazione Solo tramite cellule precursori No Si Filamenti
Scaricare