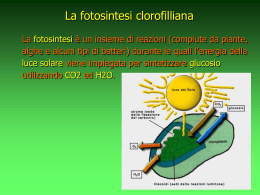
CLOROPLASTI FOTOSINTESI CLOROFILLIANA CLOROPLASTI I cloroplasti sono i plastidi contenenti la clorofilla e altri pigmenti fotosintetici necessari per la fotosintesi. Pertanto, sono organelli caratteristici della cellula vegetale e dei batteri fotosintetici. Essi possiedono una membrana esterna abbastanza permeabile, una membrana interna che rappresenta invece una barriera di permeabilità, uno spazio intermembrana ed uno spazio racchiuso dalla membrana interna chiamato stroma. Nello stroma si trova un’ulteriore membrana: la membrana tilacoidale. I tilacoidi sono impilati parallelamente a formare i cosiddetti grana. Questa membrana delimita un altro spazio all’interno del cloroplasto chiamato lume tilacoidale o spazio tilacoidale. I cloroplasti posseggono, come i mitocondri, un proprio genoma e derivano dai batteri blue-verdi. Le differenze tra cloroplasti e mitocondri sono illustrate nella prossima figura: A differenza dei mitocondri, la membrana interna dei cloroplasti non ha le creste e non contiene i complessi del trasporto elettronico. La fotosintesi avviene quindi sulla membrana tilacoidale. I cloroplasti sono ricchi di amido e di lipidi accumulati nello stroma. FOTOSINTESI Processo per mezzo del quale alcuni organismi sono in grado di catturare la luce solare e di utilizzarla per sintetizzare molecole organiche a partire da H2O e CO2, rilasciando O2. Nella biosfera gli organismi biosintetici vivono in equilibrio bilanciato con gli organismi eterotrofi. L’E solare è la fonte di tutta l’ E biologica La fotosintesi può essere aerobia ed anaerobia. Nella fotosintesi aerobia è l’H2O il donatore di elettroni e di protoni per la riduzione della CO2; non solo le piante, ma anche alcuni batteri fanno avvenire la fotosintesi però in maniera anaerobia perchè per loro l’O2 è un veleno. La fotosintesi anaerobia avviene esclusivamente da parte di alcuni batteri in cui gli elettroni non provengono dall’acqua ma da composti come ad es. H2S e quindi non producono O 2. La fotosintesi clorofilliana è un processo esclusivo delle cellule contenenti cloroplasti; avviene pertanto nelle piante (soprattutto nelle foglie ricche di cloroplasti), nelle alghe, nei batteri fotosintetici e nei cianobatteri. Le cellule fotosintetiche (autotrofe), per mezzo dell’energia solare, utilizzano l’acqua e la CO2 per produrre glucosio e O2 che vengono utilizzati dalle cellule eterotrofe (riproducendo acqua e CO2). I primi organismi fotosintetici risalgono a 33,5 miliardi di anni fa. La fotosintesi avviene in due grandi tappe: -reazioni di trasduzione dell’energia o fase luminosa (alla luce) e -reazioni di fissazione del carbonio o fase oscura (che non richiedono energia solare). La prima tappa della fotosintesi è l’assorbimento della luce da parte dei pigmenti fotosintetici: la clorofilla, una porfirina contenente Mg2+ (invece di Fe2+ come nell’Hb), che può essere distinta in clorofilla piante verdi e nei a (nelle batteri), clorofilla b (nelle piante verdi), carotenoidi (caroteni e xantofille) e ficobiline (in alghe rosse e alghe bleu-verdi): Nelle reazioni fotosintetiche di trasferimento degli elettroni o “reazioni alla luce” l’energia luminosa provoca la sintesi di ATP e la riduzione di NADP+ a NADPH, liberando O2; Nelle reazioni di fissazione del carbonio o “reazioni al buio” (perchè avvengono in assenza di luce) l’ATP e l’NADPH vengono utilizzate per trasformare la CO2 in zuccheri. La reazione di base della fotosintesi è: H2O+CO2 (CH2O)+O2 dove (CH2O) rappresenta un carboidrato: saccarosio o amido. Che la fotosintesi producesse anche O2 fu scoperto nel 1780 da un chimico inglese Priestley con un classico exp. Reazioni della fase luminosa - FOTOSISTEMI Il fotosistema II (P680) trasferisce gli elettroni dall’H2O al plastochinone, mentre il fotosistema I (P700) genera una forza riducente come NADPH. Le cellule fotosintetiche che sviluppano O2 (piante superiori e cianobatteri) contengono entrambi i fotosistemi mentre i batteri fotosintetici che non sviluppano O2 posseggono solo il fotosistema I. I due fotosistemi sono stati numerati in base alla loro scoperta. Il fotosistema II è stato scoperto dopo il fotosistema I ma il flusso di elettroni percorre prima il fotosistema II e poi il fotosistema I. Ogni fotosistema contiene da 250 a 400 molecole di pigmenti fotosintetici nelle sue due componenti: il complesso antenna e il centro di reazione. Il complesso antenna è detto così perché i pigmenti catturano l’energia solare e la trasferiscono al centro di reazione. In definitiva: l’energia luminosa, come fotoni, colpisce eccitandoli gli elettroni della clorofilla b e dei carotenoidi e viene trasferita, per risonanza, da una molecola di pigmento a quella vicina: L’elettrone della clorofilla, eccitato dalla luce, acquista energia e nel ritornare al suo stato stazionario può: 1. cedere l’energia come luce e calore; 2. trasferire l’energia per risonanza ad una molecola vicina di clorofilla e 3. il trasferimento di energia è accompagnato da un trasferimento di elettroni. Nella fotosintesi delle piante si ha il cosidetto SCHEMA Z a zigzag costituito da quattro complessi multimolecolari e da tre trasportatori singoli. I componenti della via Z si trovano nella membrana tilacoidale: Tutti i componenti, ad eccezione dei plastochinoni, sono associati a proteine: La molecola di clorofilla del P680 perdendo un elettrone si ossida caricandosi positivamente P680+. La stessa cosa succede al fotosistema I che cedendo l’elettrone alla ferredossina diventa P700+. Questo vuoto elettronico viene colmato con gli elettroni provenienti dalla scissione dell’acqua. Una volta eccitati dalla luce sono trasferiti al plastochinone QA e quindi al plastochinone QB e da quest'ultimo al pool di plastochinoni (non associati alle proteine) e quindi al citocromo b6, ai centri Ferro/Zolfo, al citocromo f e alla plastocianina (pc) (contenente rame). Dalla plastocianina gli elettroni arrivano all'accettore Ao del fotosistema I dove vengono di nuovo eccitati dalla luce e quindi passano al chinone A1 (un derivato della vitamina K) e ai centri Ferro/zolfo (Fe/S) denominati Fx, Fa e Fb. All’uscita dal fotosistema I (che si carica positivamente e diventa P700+, gli elettroni passano attraverso un altro centro Fe/S, alla ferredoxina (Fd) ed infine per mezzo del FAD (coenzima della Ferredoxina-NADP-reduttasi) arrivano all’NADPH completando il percorso della via Z. La prossima tabella riporta le differenze tra la respirazione mitocondriale ed il processo fotosintetico: Durante il passaggio degli elettroni attraverso il complesso dei citocromi b6/f, gli H+ derivanti dalla scissione dell’acqua e dai chinoni vengono pompati dallo stroma allo spazio tilacoidale generando così un gradiente elettrochimico protonico, costituito da differenza di pH (ΔpH) e da una differenza di cariche genera un potenziale di membrana (Δψ). una che Il gradiente elettrochimico protonico è la forma di energia utilizzata per fosforilare l’ADP ad ATP (attraverso il passaggio dei protoni dal lume tilacoidale allo stroma ad opera dell’enzima CFo-F1 ATP-sintasi. Il processo appena descritto è detto di fotofosforilazione non ciclica e per ogni coppia di elettroni che passa dall’acqua all’NADP+ si sintetizza 1 ATP e 1 NADPH. Poichè le reazioni di fissazione del C richiedono molto più ATP che NADPH la fotofosforilazione nei cloroplasti può passare da non ciclica a ciclica; ciò si può realizzare perchè gli elettroni provenienti dal fotosistema I anzichè andare a ridurre l’NADP+ possono essere trasmessi al complesso b6-f creando un ulteriore pompaggio di protoni nello spazio tilacoidale sintetizzando altro ATP. E quindi nel flusso ciclico è sufficiente il fotosistema I; tutto dipende dalla concentrazione di NADP+ e quindi di NADPH. Nelle piante e nei cianobatteri la sintesi di ATP e di NADPH avviene mediante un processo di fotofosforilazione non ciclica. Abbiamo visto che esistono due fotosistemi: nel fotosistema II gli atomi di O2 e di H2O si legano al Mn dell’enzima che scinde l’H2O. Quando le molecole di clorofilla del centro reattivo fotochimico vengono colpite dalla luce gli elettroni ad alta energia vengono allontanati uno per volta e appena si allontanano 4e- (il che richiede 4 protoni) l’enzima libera O2: 2H2O 4H+ +4e- +O2 Fissazione del carbonio L’energia chimica ottenuta come ATP e NADPH nella fase luminosa della fotosintesi viene impiegata nelle reazioni della fase oscura per l’organicazione della CO2 per l’ottenimento degli zuccheri e delle altre molecole organiche. La fase oscura è detta così perché non richiede la luce e avviene prevalentemente di giorno. Ciclo dei C3 o di Calvin-Benson: Sinteticamente nel ciclo dei C3 vengono utilizzati CO2, ATP e NADPH e vengono prodotti 3P-Gal (3fosfo-gliceraldeide), ADP, Pi e NADP+ Il meccanismo con cui il processo di organicazione della CO2 avviene è illustrato nella prossima figura: L’organicazione del carbonio, cioè la reazione che trasforma la CO2 in carboidrati, consiste nel far reagire la CO2 e l’H2O con ribulosio-1,5-diP per produrre 2 molecole di 3-Pglicerato ed è catalizzato dalla ribulosiobisPcarbossilasi( RuBisCO) di P.M. 5OO.OOO; questo enzima costituisce il 50% di tutte le proteine del cloroplasto, perciò è la proteina più abbondante che si conosca. Questa reazione è stata messa in evidenza da Calvin mediante autoradiogramma utilizzando CO2 radioattiva: Dalla fissazione di 3CO2 con 3 di ribulosio 1,5-diP , si ottiene un guadagno netto di una molecola di gliceraldeide-3P. Questo ciclo di fissazione del carbonio o ciclo di Calvin-Benson consuma 3ATP e 2NADPH per ogni CO2 e quindi in totale 18ATP e 12NADPH per ogni molecola di glucosio ottenuta. La gliceraldeide-3P viene trasformata in Fruttosio-6P e Glucosio-1P che reagisce con l’UTP per dare l’UDP-glucosio. Quest’ultimo reagisce con il fruttosio-6P per dare Saccarosio-P e quindi saccarosio nelle foglie. Il saccarosio rappresenta lo zucchero maggiormente utilizzato dalla pianta (ma in alcune piante può essere anche accumulato; ad es. nella barbabietola e nella canna da zucchero) che circola nei vasi del floema. Il resto della gliceraldeide-3P si trasforma nello stroma in amido che è il principale polisaccaride accumulato negli amiloplasti ( ad es. di cereali e tuberi). Il saccarosio viene secreto in grande quantità nei tubi cribrosi delle foglie ove richiama acqua per osmosi. FOTORESPIRAZIONE Quando la CO2 è presente a bassa concentrazione, al ribulosio-1,5-diP viene addizionato O2 invece di CO2 e anzichè 2 molecole di 3-Pglicerato se ne formano una di 3Pglicerato e una di Pglicolato il quale viene defosforilato a glicolato e va nei perossisomi ove due di glicolato si trasformano in una di 3-Pglicerato e 1CO2. Questo processo in cui viene consumato O2 e prodotto CO2 si chiama di fotorespirazione. Naturalmente rappresenta un inconveniente per molte piante a meno che non si verifichi un processo di adattamento come nella canna da zucchero. Ciclo dei C4 In questo ciclo la CO2 reagisce con il fosfoenolpiruvato (PEP) per formare ossalacetato e quindi malato (composto a 4 atomi di C). Il ciclo dei C4 avviene al posto del C3 in quelle cellule in cui la concentrazione di O2 è così elevata da stimolare l’attività ossigenasica del Ribulosio,1-5diP carbossilasi. Il malato prodotto diffonde verso strati cellulari in cui la concentrazione di O2 è più bassa (cellule del fascio) e si scinde in piruvato e CO2 che può entrare nel ciclo dei C3, mentre il piruvato ritorna in ciclo e si rifosforila a PEP. Il ciclo dei C4 è particolarmente presente nelle piante infestanti e in quelle che vivono in Paesi molto caldi, dove gli stomi sono quasi chiusi per evitare perdite eccessive di acqua. Genoma di cloroplasti e mitocondri La stragrande maggioranza delle proteine della cellula animale e vegetale viene sintetizzata dal genoma nucleare. Tuttavia gli organuli posseggono un proprio DNA e propri ribosomi per cui sono in grado di sintetizzare i vari tipi di RNA e di conseguenza, anche se un numero molto ridotto, di proteine. Esistono inibitori specifici delle due sintesi proteiche. Quella citosolica è inibita dalla cicloeximide mentre la sintesi proteica degli organuli no. La sintesi proteica dei mitocondri e dei cloroplasti è inibita da cloroamfenicolo, tetraciclina ed eritromicina, mentre quella citosolica non lo è. Come si sintetizzano mitocondri e cloroplasti? Solo dalla crescita e divisione di altri mitocondri e di altri cloroplasti già esistenti. Ogni organulo raddoppia la propria massa e poi si divide. Tale processo inizia con la formazione di un solco che si approfonda sempre più e la divisione avviene con il raddoppio del DNA Una caratteristica importante del genoma dei cloroplasti e dei mitocondri è che esso è costituito da DNA circolare (che nei mitocondri di cellule vegetali è molto più grande rispetto a quello dei mitocondri di cellule animali perchè contiene molti introni). Le principali differenze sono riportate in tabella: Genoma dei cloroplasti Il genoma dei cloroplasti è fortemente rassomigliante a quello dei batteri, esso contiene circa 120 geni che codificano proteine dei fotosistemi I e II, dell’ATP-sintetasi e componenti di complessi multienzimatici.
Scaricare