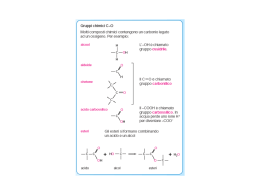
Catabolismo dei lipidi Il 90% dei lipidi è costituito da trigliceridi o triacilgliceroli o grassi neutri 5÷25 % peso corporeo dei mammiferi E’ la fonte primaria di riserva di energia degli animali Nel cuore e fegato rappresenta l’80% di apporto energetico in qualsiasi condizioni fisiologiche Sono la sola fonte di energia degli animali in letargo e degli uccelli durante la migrazione Un uomo di 70 Kg ha circa 15 Kg di trigliceridi sufficienti per 12 settimane (0,23 kg di glicogeno, sufficienti per 12 ore) I batteri non utilizzano i trigliceridi come energia di riserva Le piante producono trigliceridi e li conservano nei frutti, nelle noci e nei semi 1 Utilizzazione dei lipidi nei vertebrati Dalla biosintesi epatica dei carboidrati in eccesso Dai grassi della dieta Dalla mobilizzazione dal tessuto adiposo (adipociti) 2 Utilizzo dei lipidi dalla dieta 3 Azione dei sali biliari e delle lipasi intestinali Colesterolo Sali biliari Colesterolo 4 Struttura molecolare di un chilomicrone Le apolipoproteine sono le proteine che legano i lipidi nel plasma nella forma priva (“apo”) di lipidi Nel plasma ci sono diverse proteine e lipidi associate in strutture sferiche (lipidi al centro e proteine all’esterno) chiamate lipoproteine. Sono classificate in base alla loro densità Composizione (peso%) Lipoproteina Densità (g/ml) Proteine Fosfolipidi Colesterolo libero Esteri del colesterolo Triacilgliceroli Chilomicroni <1,006 2 9 1 3 85 VLDL 0,95-1,006 10 18 7 12 50 LDL 1,006-1,063 23 20 8 37 10 HDL 1,063-1,210 55 24 2 15 4 5 Azione della lipoproteina lipasi Chilomicroni provenienti dal sistema linfatico e flusso sanguigno La lipoproteina lipasi, attivata nei capillari dall’apoC-II, rilascia acidi grassi e glicerolo Una carenza di apoC-II comporta alti livelli di trigliceridi nel sangue 6 Mobilizzazione dei trigliceridi Gli ormoni innescano la mobilizzazione delle riserve di trigliceridi Digiuno (Glucagone), Stress (Adrenalina) Fegato 5% energia Perilipina una proteina che regola l’accesso alle gocce lipidiche Glicerolo 95% energia 7 Ingresso degli acidi grassi nei mitocondri (1) Attivazione 8 Ingresso degli acidi grassi nei mitocondri (2) Trasporto Inibita dal Malonil-CoA, primo intermedio della biosintesi degli acidi grassi 9 b-ossidazione degli acidi grassi saturi a numero pari di atomi di C (1) b-ossidazione del palmitato Elettron Tranfer Flavoprotein FADH2 FAD 10 Bilancio energetico della b-ossidazione del palmitato (1) 7 FADH2 7 NAD+ + H+ Totale fase 1 8x1 FADH2 8x3 (NAD+ + H+) 8x1 ATP Totale fase 2 Totale fase 1+2 Attivazione Bilancio finale 7x1,5=10,5 ATP 7x2,5=17,5 ATP 28,0 ATP 8x1,5=12,0 ATP 24x2,5=60,0 ATP = 8 ATP 80,0ATP 108 ATP -2 ATP 106 ATP 11 Bilancio energetico della b-ossidazione del palmitato (2) Palmitoil-CoA +23O2 +106 ADP+106Pi →CoA +106 ATP +16 CO2 + 23 H2O Resa% 2340 Kcal/mol 106 7,3Kcal/mol 100 34% 2340kcal/mol in c.s. Nelle condizioni reali intracellulari di reagenti e prodotti la resa sale oltre il 60% Gli uccelli migratori durante le transvolate utilizzano le riserve lipidiche accumulate nei giorni precedenti (i lipidi rappresentano il 70% del peso secco corporeo contro il 30% degli uccelli non migratori) Oltre alla notevole quantità di energia e alla ottima resa energetica si forma molta H2O Alcuni animali (orso bruno in letargo, cammelli) evitano la disidratazione grazie all’H2O prodotta dalla completa ossidazione dei lipidi di riserva •Cammello: 1kg di tripalmitina1,4 litri H2O •Orso bruno: perde 0,66 Kg/giorno 12 b-ossidazione degli acidi grassi insaturi a numero pari di atomi di C Insaturazione naturale in “cis” Intermedio in “trans” riconosciuto dagli enzimi della b-ossidazione 13 Ossidazione di un acido grasso polinsaturo b-ossidazione degli acidi grassi saturi a numero dispari di atomi di C b-ossidazione 5 Acetil-CoA Gli acidi grassi a numero dispari di C sono presenti nelle piante e in alcuni organismi marini. Per la loro completa ossidazione hanno bisogno di tre reazioni in più Ciclo di krebs 15 L’ossidazione degli acidi grassi è rigidamente regolata La coordinazione del metabolismo è a carico di due enzimi: Acetil-CoA Carbossilasi (ACC) primo enzima della biosintesi acidi grassi Carnitina aciltrasferasi I 16 Formazione dei corpi chetonici (1) Fegato Cervello Muscoli Cuore Surrene (Mitocondri) Steroli Citosol epatico ATP Sangue 17 Formazione dei corpi chetonici (2) Il digiuno prolungato o nel diabete mellito si ha una sovrapproduzione di acetil-CoA (blocco del ciclo di Krebs per eccessivo prelievo di ossalacetato) che porta alla formazione di corpi chetonici in eccesso (chetosi) nel sangue (chetonemia) e nelle urine (chetonuria); il conseguente abbassamento del pH (acidosi) dovuto alla presenza dei corpi chetonici acidi, stimola i reni ad espellere gli H+ in eccesso. L’escrezione di H+ è accompagnata anche dall’escrezione di Na+, K+, Pi e H2O con conseguente disidratazione (è la causa della sete eccessiva dei diabetici). 18 Le fonti di energia nel muscolo e cervello Muscolo Cervello (b-idrossibutirrato) AMP ADP Adenilato chinasi (Miochinasi) 19 Biosintesi degli acidi grassi NADPH 20 Acetil-CoA Carbossilasi (ACC):il primo enzima della sintesi degli acidi grassi Precursore principale della biosintesi La reazione avviene in due tappe come in tutte le carbossilasi biotina dipendenti (Piruvato carbossilasi, Propionil-CoA carbossilasi) 21 Acido grasso sintasi: il sitema multienzimatico della sintesi degli acidi grassi Entra una volta solo all’inizio Entra sempre ad ogni ciclo Acil Carrier Protein ACC 22 Biosintesi del palmitato (C16:0) Per produrre una molecola di palmitato occorrono 7 cicli 8 Acetil-CoA + 7 ATP + 14 NADPH + 14 H+ → Palmitato + 8 CoA + 6 H2O + 7 ADP + 7 Pi + 14 NADP+ 23 Localizzazione della biosintesi degli acidi grassi Citosol epatico: [NADPH]/[NADP+]~75 [NADH]/[NAD+]~8x 10-4 Ciò permette che il catabolismo NAD+ dipendente del glucosio (glicolisi) e la biosintesi degli acidi grassi avvenga contemporaneamente nello stesso compartimento 24 Trasporto dell’acetil-CoA dai mitocondri al citosol 25 La biosintesi degli acidi grassi è fortemente regolata Glicolisi PFK-1 Inibizione allosterica (citosolico) Attivazione allosterica (citosolico) Acil-CoA (citosolico) carnitina aciltransferasi I Acil-CoA (mitocondriale) b-ossidazione Inibizione allosterica (la fosforilazione provoca la depolimerizzazione della forma attiva dell’enzima) Regolazione covalente 26 Il palmitato è il precursore degli acidi grassi a catena lunga Sistema di allungamento degli acidi grassi (reticolo endoplasmatico liscio e mitocondri) Insaturazione degli acidi grassi (reticolo endoplasmatico liscio) Ossidasi a funzione mista Il linoleato e a-linolenato non possono essere sintetizzati dai mammiferi, ma sono prodotti dalle cellule vegetali (Acidi grassi essenziali). Una volta ingeriti possono essere convertiti in altri acidi grassi poliinsaturi tra cui l’arachidonato, precursore essenziale di lipidi regolatori (eicosanoidi) Eicosanoidi 27 Eicosanoidi Eicosanoidi Molecole segnale che agiscono nel tessuto dove sono prodotte ”ormoni ad azione locale” Arachidonato Cicloossigenasi (COX-1, COX-2) (eicosatetraenoato) via “ciclica” C20:4 (D5,8,11,14) Prostaglandine (PGF2, PGD2, PGE2, PGI2) Aspirina (TxA2) Costrizione vasi sanguigni Aggregazione delle piastrine Coagulazione del sangue Leucotrieni Costrizione vie aeree (asma) Trombossani via “lineare” Dilatazione vasi sanguigni Aggregazione piastrinica Infiammazione, Dolore Calore corporeo (febbre) Contrazione uterina Mandano in calore animali Stimolano il travaglio Secrezione mucina gastrica Causate da PG sintetizzate da COX-2 da PG sintetizzate da COX-1 28 Biosintesi Prostaglandine e Trombossani Farmaci Antinfiammatori Non Steroidei Adrenalina (ormone), Trombina (proteasi), Metillina (veleno delle api) Via ciclica Reticolo endoplasmatico Cicloossigenasi (COX-1, COX-2) Una nuova classe di FANS (Vioxx*, Celebrex), sono in grado di inibire selettivamente la COX-1 e ovviare agli effetti collaterali dei FANS di prima generazione * Ritirato per effetti collaterali al sistema cardiocircolatorio 29 Biosintesi dei lipidi NADPH 30 Biosintesi dei triacilgliceroli & fosfolipidi Fosfatidato (Diacilglicerolo 3.fosfato) precursore comune 31 Controllo ormonale della biosintesi dei trigliceridi Citrato liasi Complesso piruvato deidrogenasi Perdita di peso 32 Ciclo del triacilglicerolo Durante il digiuno, negli adipociti dei mammiferi, gli acidi grassi ottenuti dall’idrolisi dei trigliceridi, sono in parte esportati tramite il sangue per essere utilizzati come fonte di energia (es: ai muscoli), il resto va al fegato dove sono convertiti in trigliceridi.Questi ritornano agli adipociti come acidi grassi, per idrolisi di una lipoproteina lipasi, e riconvertiti in trigliceridi. Questo “ciclo del triacilglicerolo” ha la funzione di mantenere una concentrazione costante di acidi grassi liberi nel sangue come fonte di energia immediatamente utilizzabile in caso di emergenza. 33 Regolazione del ciclo del triacilglicerolo Il ciclo del triacilglicerolo è controllato dai glucocorticoidi del surrene (es: cortisolo) attraverso la regolazione dell’attività della PEPCK Gliceroneogenesi La presenza dell’ormone, indotta dal digiuno, stimola la gliceroneogenesi nel fegato e la reprime nel tessuto adiposo aumentando i livelli di acidi grassi liberi nel sangue; ciò può interferire con l’utilizzo del glucosio nel muscolo e promuovere la resistenza all’insulina (diabete tipo 2). I tiazolidinedioni, inducendo i livelli di PEPK, sono in grado di curare questa patologia. 34 Biosintesi del colesterolo Nei vertebrati la maggior quantità di colesterolo viene sintetizzato nel citosol del fegato in quattro tappe a partire dall’acetil-CoA Le prime due reazioni della prima tappa sono identiche alla formazione dei corpi chetonici (nei mitocondri epatici) 6 unità isopreniche 35 Funzioni del colesterolo CH3 Nei vertebrati il colesterolo, prodotto principalmente nel fegato, viene : incorporato nelle membrane degli epatociti (una piccola parte) utilizzato nella sintesi di acidi biliari per la digestione dei lipidi esportato in altri organi, in forma libera o esterificato, tramite le proteine plasmatiche, per la produzione degli ormoni stereoidei e vitamina D H C (CH2)3 CH3 CH3 CH CH3 CH3 HO 36 Trasporto del colesterolo e altri lipidi (1) Il colesterolo, gli esteri del colesterolo, i triacilgliceroli e i fosfolipidi, per essere trasportati da un tessuto ad un altro tramite il plasma, essendo insolubili in acqua, devono essere inglobati in specifiche lipoproteine plasmatiche Composizione (peso%) Lipoproteina Densità (g/ml) Proteine Fosfolipidi Colesterolo libero Esteri del colesterolo Triacilgliceroli Chilomicroni <1,006 2 9 1 3 85 VLDL 0,95-1,006 10 18 7 12 50 LDL 1,006-1,063 23 20 8 37 10 HDL 1,063-1,210 55 24 2 15 4 37 Trasporto del colesterolo e altri lipidi (2) Colesterolo “cattivo” associato alle LDL Colesterolo “buono” associato alle HDL Il colesterolo LDL in eccesso viene rimosso dalle HDL, altrimenti si deposita sulle pareti delle arterie (aterosclerosi) Eccesso di lipidi e carboidrati nella dieta Utilizzati come fonte di energia Riserva sotto forma di gocce lipidiche 38 HDL: gli spazzini del colesterolo “cattivo” Le HDL hanno origine nel fegato e intestino sotto forma di piccole particelle a disco piatto; contengono molte proteine e poco colesterolo in forma non esterificata. La presenza della lecitina-colesterolo aciltransferasi (LCAT) sulla superficie delle HDL esterifica il colesterolo dei chilomicroni e delle rimanenze di VLDL, che entra all’interno delle HDL (struttura sferica matura). La particella ricca di colesterolo ritorna al fegato, dove viene scaricato il colesterolo 39 Assunzione di colesterolo “cattivo” tramite endocitosi mediata da recettore Il colesterolo che entra nella cellula tramite questa via può essere incorporato nelle membrane o esterificato dalla Acil-CoA-colesterolo aciltransferasi (ACAT) per essere conservato in gocce lipidiche 40 Regolazione della sintesi del colesterolo Quando la concentrazione del colesterolo sintetizzato e di quello introdotto con la dieta supera la quantità necessaria per la sintesi delle membrane, sali biliari e steroli, si accumula nei vasi sanguigni e può causare placche con ostruzione dei vasi (aterosclerosi) Alcuni inibitori competitivi della HGM-CoA reduttasi si sono rivelati efficaci nell’abbassare i livelli di colesterolo LDL ematico 41 Metabolismo degli acidi grassi nel fegato , fosfolipidi di membrana VLDL Adipociti (riserva) Cuore,Muscolo Albumina (fino a 10 per molecola) (energia) Tessuti Integrato nella membrana (energia: 1/3 del fabbisogno del cuore fino a 2/3 nel cervello) 42
Scaricare