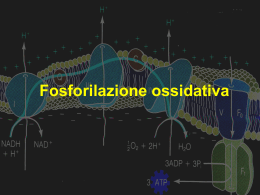
Reazioni redox ØLe reazioni redox possono produrre lavoro biologico ∆E = Eossidante - Eriducente ∆G = -nF∆E o ∆G’ = o -nF∆E’ Funzioni della catena respiratoria La catena respiratoria è parte essenziale del processo di fosforilazione ossidativa. Quest’ultimo processo fornisce alla maggior parte delle cellule l’energia (ATP) necessaria per tutti i tipi di lavoro: meccanico, osmotico, biosintetico etc. La catena respiratoria determina: Ø la riossidazione del NADH a NAD e del FADH2 a FAD; Ø La riduzione dell’ossigeno (O2) che viene trasformato in H2O; Ø La generazione di un potenziale energetico di natura elettrochimica, necessario per la fosforilazione dell’ADP da parte del fosfato inorganico (Pi), per formare l’ATP. Mitocondrio Catena Respiratoria Localizzazione cellulare: membrana mitocondriale interna La catena respiratoria è costituita da una serie di reazioni redox tra loro concatenate in modo similare a quello con cui sono concatenate le reazioni di una via metabolica. Cioè il prodotto della prima reazione (la sostanza che riceve gli elettroni dai coenzimi NADH o FADH2) li cede al substrato ossidato della seconda reazione e così via finché gli elettroni verranno raccolti dall’ossigeno molecolare (O2), substrato ossidante dell’ultima reazione redox. Tutte le molecole trasportatori di elettroni che partecipano alla catena respiratoria possono esistere in due stati: uno ossidato ed uno ridotto (coppie redox). Nella catena respiratoria tali coppie sono ordinate secondo un potenziale redox crescente (potenziale di riduzione). All’inizio della catena respiratoria viene posta la coppia redox NAD+/ NADH, ed alla fine la coppia redox O2/ H2O. L’ossigeno diffonde infatti nei mitocondri dove viene utilizzato nella fosforilazione ossidativa per costruire l’ATP. Anche per l’ossigeno esistono due forme importanti per la catena respiratoria, una ossidata (O2) ed una ridotta O2- che viene prodotta quando 2 elettroni si legano ad ogni atomo di ossigeno della molecola O2 (totale 4 elettroni). Questi ioni formano H2O combinandosi ognuno con 2 protoni (H+): O2 + 4e2O2- + 4H+ 2O22H2O ØLa catena respiratoria è catalizzata da tre complessi principali (complesso-I, complesso-III e complesso-IV) formati ognuno da diverse proteine (tra cui alcuni citocromi), legate tra loro in una sorta di struttura quaternaria, e da molecole non proteiche quali i gruppi prostetici FMN, eme, “centri ferro-zolfo” di differenti strutture, ioni rame. Oltre a questi tre complessi c’è anche un quarto complesso (complesso-II) necessario per fornire alla catena principale gli elettroni ceduti dal coenzima FADH2. ØI centri ferro-zolfo contengono ioni ferro legati sia a zolfo inorganico che a gruppi –SH dell’ammino acido cisteina, gruppi presenti nelle catene laterali di proteine dei complessi. ØTutti questi complessi sono localizzati nella membrana interna del mitocondrio. In tale membrana essi sono fisicamente separati; tuttavia sono collegati tra loro da altre molecole (il coenzima-Q ed il citocromo-C) che trasportano elettroni da un complesso all’altro. Schema della catena respiratoria. Complesso-I = NADH-CoQ reduttasi Complesso-III = citocromo-C reduttasi Complesso-IV = citocromo-C ossidasi Complesso-II = succinato deidrogenasi La catena respiratoria è come una cascata termodinamicamente spontanea che, salto dopo salto, trasferisce elettoni all’ossigeno molecolare trasformandolo in acqua. Come una cascata di un fiume può convertire energia potenziale in altra forma di energia durante la caduta, così la cascata di elettroni (catena respiratoria) converte l’energia del NADH e del FADH2 in altre forme di energia (potenziale elettrochimico) che, a sua volta, sarà convertita nell’energia chimica dell’ATP durante il processo della fosforilazione ossidativa. Trasporto degli elettroni nel complesso-I (NADH-CoQ reduttasi) e nel complesso-III (citocromo-c reduttasi). I citocromi b562 (bL bL) e b566 (bH bH) fanno parte del complesso-III. Nel complesso-I il NADH prodotto nelle vie cataboliche viene ossidato trasformandosi in NAD+. Quest’ultima forma del coenzima è necessaria per il buon funzionamento delle vie cataboliche (glicolisi, β-ossidazione, ciclo di Krebs etc.) e perciò viene riciclata. Contemporaneamente, due elettroni del NADH vengono ceduti al flavinmononucleotide (FMN), che a sua volta li cede, uno alla volta, ai centri ferro-zolfo. Quindi, un centro ferro-zolfo ridotto (Fe2+-S) li cede, ancora uno alla volta, al CoQ (ubichinone) che viene trasformato in CoQH2, (CoQ ridotto o ubichinolo). Nel complesso-III, il coenzima-QH2 cede elettroni, uno alla volta, al complesso ferro-zolfo (Fe-S) che a sua volta li cede al citocromo-c1, il quale a sua volta li cede al citocromo-c[Fe3+] trasformandolo nella forma ridotta (citocromo-c[Fe2+]. Come si vede nello schema, solo la forma QH2 del coenzima può cedere un elettrone al centro Fe-S, trasformandosi in QH• (forma questa detta semichinone). Una volta formate 2 molecole di QH•, queste vengono trasformate: una nella forma ossidata e l’altra nella forma ridotta dai citocromi b566 (bH) e b562 (bL) contenuti nel complesso-III: QH• + QH• Q + QH2 Mentre Q (forma ossidata) può tornare a ricevere altri elettroni dal complesso-I, QH2 cede di nuovo elettroni al complesso-III. Nel complessocomplesso-IV avviene la riduzione dell’ossigeno (O2) ad acqua. Infatti, lo schema sotto riportato mostra come un totale di 4 elettroni + 4 ioni H+ vengono ceduti al complesso IV, perché questo li possa combinare con una molecola di O2 allo scopo di formare 2 molecole di H2O. E’ essenziale che O2 riceva tutti i 4 elettroni: infatti, se ne ricevesse solo uno, si formerebbe l’ione superossido (O2-), una sostanza pericolosa per la cellula, perché è una forma radicalica molto reattiva. Anche se O2 ricevesse solo 2 elettroni, si formerebbe un’altra sostanza pericolosa, lo ione perossido (-O-O-) Trasporto degli elettroni nel complesso-IV (Citocromo-c ossidasi) Il complesso IV, olre a numerose catene polipeptidice, contiene due gruppi eme (eme-A ed eme A3) e due centri binucleari formati da ioni rame (Cu-A e Cu-B). Trasporto degli elettroni nel complesso-IV Ø1° stadio: stadio la molecola O2 si lega all’ione ferroso (Fe2+) dell’eme-A3 e viene rapidamente ridotta a ione perossido (O22-) acquistando 2 elettroni: uno dal Fe2+ dell’eme ( Fe2+ Fe3+ + e- ) e l’altro dal vicino ione Cu+ (Cu+ Cu2+ + e- ). Come si vede, in questa fase sia il ferro sia il rame vengono ossidati dall’ossigeno che accetta i due elettroni ceduti dagli ioni metallici. Ø2° stadio: stadio ora inizia l’acquisizione dei 4 elettroni ceduti al complesso dal citocromo-c ridotto. Il primo elettrone + 2 H+, insieme alla ulteriore ossidazione dell’ione ferrico (Fe3+) a ione ferrile (Fe4+), causano la rottura del legame tra i due atomi di ossigeno della molecola O2: si forma così la prima molecola di H2O (che resta legata a Cu2+) e di un ione O2-, che resta legato all’ione Fe4+. Ø3° stadio: il secondo elettrone determina la riduzione di Fe4+ a Fe3+, ed i due ioni H+ che entrano nel complesso insieme all’elettrone determinano la formazione della seconda molecola di H2O. Ambedue le molecole di acqua vengono rilasciate dal complesso-IV. Ø4° stadio: stadio entra il terzo elettrone e riduce lo ione rameico (Cu2+) a ione rameoso (Cu+). Ø5° stadio: stadio entra il quarto elettrone e riduce lo ione ferrico (Fe3+) a ione ferroso (Fe2+), ripristinando così la forma iniziale del complesso-IV, già pronta a ricominciare un nuovo ciclo di reazioni per ridurre ad acqua una seconda molecola di O2. Schema della catena respiratoria che mette anche in evidenza il ruolo attivo dei complessi I, III e IV nella esportazione di ioni H+ dalla matrice allo spazio tra le due membrane mitocondriali. Come si vede, il complesso-II non pompa nessun ione H+ attraverso la membrana: il suo ruolo è quello di trasferire elettroni dal FAD al CoQ. Teoria Chemio-Osmotica di Mitchell (premio Nobel) Il gradiente elettroelettro-chimico ØDurante il trasferimento degli elettroni dai coenzimi NADH e FADH2 all’ossigeno avviene, in maniera strettamente accoppiata a tale trasferimento, un fenomeno molto importante per la generazione mitocondriale dell’ATP, la formazione di un gradiente energetico di natura elettro-chimica. ØTre dei complessi della catena respiratoria (I, III, IV), oltre a trasportare gli elettroni all’ossigeno, trasferiscono ognuno un certo numero di protoni (H+) dall’interno del mitocondrio (matrice) all’esterno (nel citoplasma). Quindi, i complessi sono anche pompe protoniche direzionali che determinano la formazione di un gradiente di pH tra il compartimento interno e quello esterno del mitocondrio (cioè, il pH del fluido contenuto dentro il mitocondrio diviene più basico, mentre il pH del fluido che si trova nello spazio intermembrana diviene più acido). ØInoltre, dato che i protoni hanno una carica ionica positiva, tale trasferimento provoca sia una diminuzione della carica elettrica positiva sul lato interno della membrana mitocondriale sia un aumento della carica positiva sul lato esterno. Ciò determina una differenza di potenziale elettrico tra le due facce della membrana mitocondriale interna. Il potenziale di membrana ed il gradiente di pH costituiscono insieme un potenziale elettro-chimico (una forma di energia che i mitocondri utilizzano per costruire ATP a partire da ADP e fosfato inorganico (Pi). Il complesso della ATP sintasi (Fo(Fo-F1) ed il meccanismo chemioosmotico di formazione dell’ dell’ATP. La reazione di formazione dell’ATP a partire da ADP e Pi è la seguente: Adenosina-P∼P + Pi ADP Adenosina-P∼P∼P P ATP Il simbolo “ ∼ “ viene utilizzato per indicare legami ricchi di energia Questa reazione avviene, in condizioni standard, solo se viene fornita un’energia libera di Gibbs di 7,3 kcal/mole di ATP formato (30,5 kJ/mole), appunto l’energia che il potenziale elettrochimico generato dai complessi I. III e IV della catena respiratoria possono fornire. L’enzima che catalizza la reazione di sintesi dell’ATP è l’ ATPATP-sintasi . Si trova anch’esso intessuto nella membrana mitocondriale interna. Esso è costituito da due parti con funzioni differenti: la prima detta Fo è costituita da un complesso di proteine che formano un canale per far passare gli ioni H+ attraverso la membrana mitocondriale interna, mentre la seconda, detta F1, di forma sferoidale, è formata anch’essa da un complesso di diverse proteine e contiene i siti attivi dell’enzima, che sono tre, e si trovano in ogni istante in uno stato funzionale differente. Uno dei tre siti lega fortemente l’ATP (sito T), il secondo (sito O) è vuoto ed a bassa affinità, mentre il terzo contiene ADP e Pi (sito L). Quando passano gli ioni H+ (protoni) attraverso la membrana in direzione citoplasma ? mitocondrio, il sito che contiene l’ATP si apre e rilascia la molecola di ATP nella matrice, perché la sua affinità diminuisce dato che i tre siti cambiano la loro forma in modo sincrono, con conseguente cambiamento del loro stato funzionale. Il sito T diviene O, il sito O diviene L (e lega nuovamente ADP e Pi), mentre il sito L diviene T (cioè cataliticamente attivo, e con alta affinità per l’ATP); perciò capace di legare tra loro ADP e Pi formando nuovo ATP. Ciò significa che la dissipazione del gradiente energetico provocata dal ritorno dei protoni all’interno del mitocondrio viene utilizzata dal complesso Fo-F1 della ATP-sintasi per produrre l’ATP. Questo processo biologico, che accoppia le reazioni redox della catena respiratoria con la fosforilazione dell’’ADP, è estremamente importante per la vita di molti tipi di cellule (aerobiotiche), e viene indicato complessivamente come FOSFORILAZIONE OSSIDATIVA ∆G = 20 kJ/mole kJ/mole di ioni H+ Dato che per ogni NADH ossidato dalla catena respiratoria ci sono sono 10 e- pompati nello spazio intermembrana, 200 kJ dei 220 disponibili dalla ossidazione del NADH sono conservati dal potenziale elettrochimico. Sostanze disaccoppianti
Scaricare