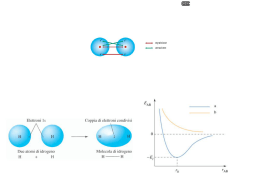
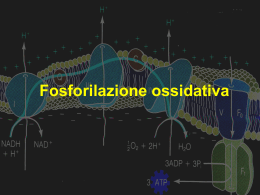
I mitocondri sono gli organuli responsabili della produzione di energia necessaria alla cellula per crescere e riprodursi. Queste reazioni, che nel loro insieme costituiscono il processo di "respirazione cellulare", comportano il consumo di ossigeno e la produzione di anidride carbonica. Infatti il glucosio viene bruciato in presenza di ossigeno e l'energia liberata, immagazzinata sotto forma di ATP Organuli di forma bastoncellare. O.5 - 1 μm di spessore Fino a 10 μm di lunghezza. Elevato numero per cellula. La struttura base dei mitocondri “animali” e “vegetali” e’ simile con una matrice interna, una membrana interna, uno spazio intermembrana e una membrana esterna. Distribuzione dei Mitocondri Tutte le cellule hanno mitocondri. La maggior parte dei mitocondri è localizzata nella porzione della cellula che ha maggior richiesta energetica. Meno abbondanti in quelle cellule che lavorano in condizioni anaerobiche. OGGI SAPPIAMO CHE IL MITOCONDRIO è DELIMITATO DA DUE MEMBRANE ALTAMENTE SPECIALIZZATE CHE HANNO FUNZIONI DIVERSE E CHE INSIEME CREANO DUE COMPARTIMENTI MITOCONDRAILI SEPARATI: Doppia membrana: – Membrana esterna Liscia – Membrana interna ripiegata: Creste mitocondriali MEMBRANA ESTERNA : ha funzioni protettive, essa conferisce resistenza all’organello. Risulta caratterizzata dalla presenza di una grossa proteina che forma canali (detta porina),che la rende permeabile alle molecole di dimensioni inferiori ai 5000 dalton. SPAZIO INTERMEMBRANA: detto spazio bianco che risulta essere chimicamente equivalente al citosol, per quel che riguarda le piccole molecole che contiene. LA MEMBRANA INTERNA: ha proprietà di permeabiltà selettiva, grazie alla presenza di specifici trasportatori, si invagina a formare dei ripiegamenti di forma tubulare detti “creste”. Queste introflessioni sono strettamente legate alla funzione del mitocondrio, in quanto la membrana interna ospita i complessi respiratori di trasporti di elettroni,i traslocatori di metaboliti e l’ATP sintasi. Le creste inoltre facilitano un intimo contatto con la: MATRICE: in essa sono presenti gli enzimi per l’ossidazione del piruvato e quelli per il ciclo dell’acido citrico. Inoltre contiene diverse copie di DNA mitocondriale, ribosomi e tRNA mitocondriali . Le creste differiscono in lunghezza,forma e numero, a seconda delle richieste energetiche della cellula. Cellule normali:creste si allungano per metà della matrice. Le creste sono presenti in numero elevato in corrispondenza di una elevata richiesta energetica. Ad esempio nelle cellule muscolari le creste attraversano tutta la matrice e si presentano impacchettate molto strette. Mitocondri “Classici”Creste a ripiani. Metà della matrice. Mitocondri Attivi Creste a ripiani strettamente impilate Attraverso tutta la matrice Le membrane mitocondriali sono composte principalmente da proteine, lipidi in misura minore. Le proteine mitocondriali sono costantemente rinnovate. Membrana esterna Ricca di proteine Multipasso,dette Porine. Ricordano le proteine delle membrane batteriche. Teoria del simbionte. Membrana interna Proteine dei complessi respiratori I mitocondri sono stati essenziali per l’evoluzione degli organismi eucarioti. Senza di essi gli organismi superiori sarebbero dipendenti dalla glicolisi anaerobia per tutto il loro ATP. NEI MITOCONDRI IL METABOLISMO DEGLI ZUCCHERI è COMPLETATO : IL PIRUVATO, E’ IMPORTATO NEL MITOCONDRIO E OSSIDATO COMPLETAMENTE DA O2 a CO2 e H2O. QUESTO COMPORTA UNA RESA ENERGETICA NOTEVOLMENTE SUPERIORE RISPETTO ALLA SOLA GLICOLISI. Fasi in cui in cui si puo’suddividere la respirazione: 1. Glicolisi 2. Ciclo degli acidi tricarbossilici 3. Catena di trasporto degli elettroni mitocondriale La respirazione e’ un processo multistep, in cui il glucosio viene completamente ossidato a CO2 e H2O … con concomitante produzione di ATP . 3atomi di C Alla fine del ciclo si riottiene l’ossalacetato che puo’ rientrare nel ciclo . 2atomi di C Viene legato all’ossalacetato 4C dando: Il citrato va incontro ad una serie progressiva di ossidazioni Le molecole combustibili sono trasportate attraverso la membrana mitocondriale interna nella MATRICE, dove sono poi convertite nell’intermedio metabolico cruciale l’ACETIL CoA. Questo entra quindi nel ciclo di Krebs,dove gli atomi di carbonio dell’acetil CoA vengono convertiti in CO2 che è poi rilasciato dalla cellula come prodotto di rifiuto. Ma la cosa più importante è che il ciclo genera elettroni ad alta energia, portati dalle molecole trasportatrici attivate “NADH e FADH2“. QUESTI ELETTRONI SONO POI TRASFERITI DALLA MATRICE-> ALLA MEMBRANA INTERNA, dove entrano nella catena di trasporto degli elettroni. La respirazione cellulare si svolge nei mitocondri delle cellule eucariotiche o nel periplasma dei batteri; in questa reazione si ossidano i composti del carbonio producendo anidride carbonica e acqua e contemporaneamente si genera energia. Nel corso delle reazioni di ossidoriduzione biologiche gli elettroni non sono lasciati liberi nel solvente, ma vengono trasferiti a coenzimi, quali il NAD+ (Nicotinamide Adenina Dinucleotide) o il FAD (Flavin Adenina Dinucleotide), definiti appunto trasportatori intermedi di elettroni, che portano gli elettroni alla catena respiratoria mitocondriale. Il bilancio ossidoriduttivo nelle cellule può essere valutato appunto dal rapporto tra NAD+ (forma ossidata) NADH (forma ridotta). Questo valore controlla la interconversione delle molecole organiche presenti nella cellula e può essere alterato in molte situazioni patologiche, come ad esempio ipossia (scarsità di ossigeno), shock o sepsi. Teoria chemiosmotica Il NAD+ e il FAD trasportano gli elettroni ai complessi proteici situati sulla membrana interna dei mitocondri, che nel loro insieme formano la catena respiratoria. L’ordine con cui i complessi della catena respiratoria accettano gli elettroni non è casuale, ma segue il loro potenziale di ossidoriduzione, dal più negativo (complesso I) al più positivo (complesso IV) fino all’ossigeno molecolare che è l’accettore ultimo. In alcune tappe del trasferimento elettronico l’energia libera prodotta è sufficiente per trasportare protoni (H+) dalla matrice mitocondriale allo spazio intermembrana. Si origina così un gradiente elettrochimico di protoni; l’energia potenziale contenuta nel gradiente protonico viene sfruttata per compiere un lavoro biologico, e cioè la sintesi di ATP . L’accoppiamento tra il trasporto degli elettroni, il trasporto dei protoni e la sintesi di ATP è appunto alla base della teoria chemiosmotica (1961) formulata da Peter Mitchell (Premio Nobel nel 1978). Sulla membrana interna del mitocondrio si trovano particolari molecole, capaci di accettare e donare elettroni e perciò dette trasportatori (citocromi, flavoproteine e coenzima Q). A queste il NADH e il FADH2, derivanti dal ciclo di Krebs, cedono i propri elettroni, convertendosi nella forma ossidata e potendo quindi essere riutilizzati in un nuovo ciclo. Il trasferimento di elettroni che si attiva lungo i trasportatori è un processo che libera energia, ed è responsabile della trasformazione dell'O2 (l'accettore finale degli elettroni, proveniente dalla respirazione) ad acqua, e della alta resa energetica della respirazione cellulare, in termini di sintesi di molecole di ATP. Questa energia, infatti consente il pompaggio di ioni idrogeno H+ dalla matrice verso lo spazio tra le due membrane mitocondriali, e l'aumento della loro concentrazione in questa sede. Per il fenomeno della chemiosmosi, questi ritornano verso la matrice passando attraverso speciali canali proteici, detti ATP-sintetasi; tale flusso di ioni mette a disposizione energia per la sintesi di ATP. Catena di trasporto degli elettroni Catalizza un flusso di elettroni dal NADH (FADH2) all’O2 (accettore finale degli elettroni nel processo respiratorio) Consiste di un gruppo di proteine trasportatrici di elettroni legate alla membrana interna mitocondriale. La perdita degli elettroni da parte di NADH e FADH2 rigenera anche NAD+ e FAD, che sono necessari per un metabolismo ossidativo continuo. Gli elettroni vengono trasferiti lungo la catena di trasporto degli elettroni, fino ad arrivare all’accettore finale: O2. Il complesso I accetta elettroni da NADH e li trasferisce all’ubichinone che a sua volta li trasferisce al complesso dei citocromi b-c1 (complesso III). Il complesso accetta elettroni dall’ubichinone e li passa al citocromo c, che trasporta il suo elettrone al complesso della citocromo ossidasi (complesso IV). Questo accetta un elettrone alla volta dal citocromo c e ne passa 4 alla volta dell’O2. Infatti occorrono 4 elettroni alla molecola di O2 per produrre acqua. Durante il trasferimento di elettroni l’ubichinone e il citocromo c servono da trasportatori mobili che portano gli elettroni da un complesso al successivo. Il trasferimento di elettroni è accoppiato al rilascio orientato di H+ attraverso la membrana interna. I protoni sono pompati attraverso la membrana da ciascun complesso enzimatico respiratorio. Questo movimento di protoni ha due conseguenze principali: 1) genera un gradiente di pH attraverso la membrana mitocondriale interna, con il pH più alto nella matrice rispetto al citosol; 2)genera un gradiente di voltaggio attraverso la membrana interna, con l’interno negativo e l’esterno positivo. Il gradiente di pH e quello di voltaggio, insieme, costituiscono il GRADIENTE PROTONICO ELETTROCHIMICO. Questo gradiente viene usato per spingere la sintesi di ATP nel processo cruciale della fosforilazione ossidativa. Ciò è reso possibile dall’enzima ATP SINTASI. ATP SINTASI L’enzima è composto da una porzione di testa, chiamata F1, e un trasportatore transmembrana di protoni, chiamato F0. Sia F1 che F0 sono formati da subunità multiple Questo enzima crea una via idrofilica attraverso la membrana interna che permette ai protoni di scorrere lungo il loro gradiente elettrochimico. Non appena questi ioni passano attraverso l’ATP sintasi, vengono usati per spingere la reazione energeticamente sfavorevole fra ADP E Pi che produce ATP. In assenza di mitocondri molti L’ATP è un composto chimico che può organismi cedere energia e viene quindi utilizzata eucarioti non dalla cellula come "Gettone energetico" sarebbero in in tutte le reazioni cellulari. grado di utilizzare Diversamente dagli organismi aerobi, che non possono vivere in assenza di ossigeno, gli organismi anaerobi prosperano anche, o solo, in assenza di questo gas e le loro cellule sono prive di mitocondri. l'ossigeno per ricavare dagli alimenti tutta l'energia necessaria alle loro funzioni vitali. NEL CITOSOL(glicolisi): 1glucosio 2piruvato+ 2 NADH +2 ATP NEL MITOCONDRIO (piruvato deidr.e ciclo dell’a.citrico): 2piruvato 2 acetil CoA+ 2NADH 2acetil Coa TOT: glicolisi + CICLO Krebs 6 NADH + 2 FADH2 +2 GTP 10NADH + 2 FADH2 + 2GTP* *Il guanosintrifosfato è coinvolto nel trasferimento di energia entro la cellula: per ogni ripetizione del ciclo di Krebs, infatti, viene generata una molecola di GTP, equivalente alla formazione di una molecola di ATP, poiché il GTP viene subito convertito in ATP. I MITOCONDRI HANNO UN PROPRIO DNA DI FORMA CIRCOLARE E RIBOSOMI DI PICCOLE DIMENSIONI. IN REALTA’ IL SUO GENOMA E’ MOLTO PICCOLO. I mitocondri sono dotati di un proprio DNA (mtDNA). L’mtDNA è estremamente piccolo, ha struttura circolare, e contiene solamente 37 geni: 13 geni che codificano alcune delle subunità proteiche dei complessi della catena respiratoria e 24 geni che codificano molecole indispensabili alla sintesi di tali subunità (2 RNA ribosomali, rRNA e 22 RNA transfer, tRNA). Durante la fecondazione, i mitocondri presenti nel nuovo individuo (zigote) provengono solo dalla cellula uovo (oocita). Una madre portatrice di una mutazione del mtDNA trasmetterà tale mutazione a tutti i propri figli, ma solo le figlie trasmetteranno a loro volta la mutazione alla loro progenie (eredità matrilineare). Mutazioni deleterie del mtDNA generalmente colpiscono solo alcuni genomi mitocondriali; perciò, all’interno di ogni cellula e, conseguentemente, in ogni tessuto sono presenti due popolazioni miste di mtDNA: una normale ed una mutata (eteroplasmia). Al contrario dei geni nucleari che sono presenti nell’uomo in duplice copia (allele materno ed allele paterno), vi sono centinaia di molecole di mtDNA all’interno di ogni cellula. In un individuo normale, tutte le molecole di mtDNA sono identiche (omoplasmia). Solo quando il numero di genomi mutati supera un valore critico (effetto soglia) si inizia ad avere una riduzione dell’attività OXPHOS e la conseguente comparsa di manifestazioni cliniche di malattia. http://www.mitopedia.org/patologie.php Probabilmente quasi due miliardi di anni fa un batterio aerobio è stato fagocitato da una cellula eucariotica primitiva, stabilendosi nel suo citoplasma. Si serebbe così creato un rapporto simbiotico tra i due microorganismi: il batterio forniva energia in cambio di un ambiente stabile e riparato e di un apporto rapido di sostanze nutritive. Questo batterio sarebbe stato l’antenato dell’attuale mitocondrio. Origine della cellula eucariote e dei mitocondri In due step successivi simultanea (hydrogen hypothesis)
Scaricare