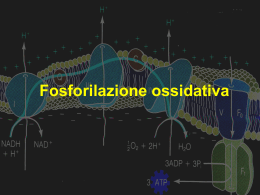
Nella fosforilazione ossidativa il potenziale di trasferimento degli elettroni del NADH e del FADH2 viene convertito nel potenziale di trasferimento del gruppo fosforico dell’ATP. Quando due coppie redox coniugate sono presenti nella stessa soluzione, il trasferimento di elettroni dal donatore di elettroni di una coppia all’accettore di elettroni dell’altra coppia dipende dalle affinità relative dell’accettore di elettroni di ciascuna coppia. Il potenziale di riduzione standard (E0) è una misura (in volt) di questa affinità e viene determinato in un esperimento di questo tipo: Gli elettrochimici hanno scelto come standard di riferimento la reazione: H+ + e- Æ 1/2 H2 L’elettrodo su cui ha luogo questa semireazione (detto semicella) viene arbitrariamento considerato con un potenziale di riduzione pari a 0,00 V. Quando l’elettrodo ad idrogeno viene collegato a un’altra semi-cella in cui le specie ossidata e ridotta sono presenti in condizioni standard (1 M o 1 atm), gli elettroni tenderanno a fluire verso la semicella con il potenziale redox più elevato. Per convenzione la semi-cella con la tendenza più forte ad acquistare gli elettroni viene indicata con un valore (in volt) di E0 positivo. Come per la definizione di ΔG’0, i biochimici definiscono lo stato standard per le reazioni di ossido-riduzione a pH 7,0, ed esprimono il potenziale sotto forma di E’0, il potenziale di riduzione standard a pH 7. Un potenziale di riduzione standard negativo indica che una sostanza ha un’affinità per gli elettroni più bassa di quella dell’H2. Un potenziale di riduzione standard positivo indica che una sostanza ha un’affinità per gli elettroni più alta di quella dell’H2. La variazione di energia libera standard è correlata alla variazione nel potenziale di riduzione dalla relazione: ΔG’0= -nFΔE’0 n = numero di elettroni trasferiti; F = variazione di energia libera generata da 1 mole di elettroni e da un cambio di potenziale di 1 volt. La differenza di potenziale di riduzione (ΔE’0) nella catena respiratoria è di 1,14 V a) b) 1/2 O2 + 2 H+ + 2 e- Æ H2O NAD+ + H+ + 2 e- Æ NADH quindi: c) 1/2 O2 + NADH + H+ Æ H2O + NAD+ E’0 = + 0,82 V E’0 = - 0,32 V ΔE’0 = + 1,14 V L’energia libera per questa reazione è ΔG’0= -nFΔE’0= -52,6 kcal/mole Un ΔE’0 positivo, come un ΔG’0 negativo, identificano una reazione esoergonica in condizioni standard. Schema essenziale della fosforilazione ossidativa L’ossidazione e la sintesi di ATP sono accoppiate mediante flussi protonici transmembrana. I mitocondri (in verde) formano una rete all’interno di un fibroblasto I mitocondri ossidano i combustibili carboniosi per produrre energia cellulare. Questa trasformazione richiede il trasferimento degli elettroni attraverso grandi complessi proteici, alcuni dei quali pompano protoni, generando un gradiente protonico che favorisce la sintesi di ATP. Gli elettroni sono incanalati verso accettori universali di elettroni La fosforilazione ossidativa ha inizio con l’ingresso degli elettroni nella catena respiratoria. La maggior parte di questi elettroni è il frutto dell’azione di deidrogenasi che li raccolgono dai processi catabolici per poi incanalarli verso accettori universali di elettoni: i nucleotidi piridinici (NAD+ o NADP+) o flavinici (FMN o FAD) Gli elettroni passano attraverso una serie di trasportatori legati alla membrana Ubichinone Anche detto coenzima Q è un benzochinone con una catena laterale isoprenoide molto lunga. L’ubichinone è idrofobico e quindi liberamente diffusibile nel doppio strato lipidico della membrana mitocondriale interna. In questo modo può agire da ponte fra trasportatori di elettroni meno mobili nella membrana. I citocromi sono proteine contenenti ferro Il gruppo prostetico dei citocromi è il gruppo eme. I citocromi sono proteine con un caratteristico elevato assorbimento della luce nel campo visibile dovuto al gruppo prostetico eme, contenente ferro Spettri di assorbimento del citocromo c nella forma ossidata (in rosso) e nella forma ridotta (in blu). Le proteine ferro-zolfo Nelle proteine ferro-zolfo il ferro è associato ad atomi di zolfo inorganico e/o ad atomi di zolfo di residui di cisteina della proteina (centri Fe-S) Il potenziale di riduzione standard del ferro nei centri Fe-S dipende dal tipo di centro e dalle specifiche interazioni che ha con la proteina in cui è inserito. Centro Fe-S semplice Un solo ione Fe è circondato da 4 residui di cisteina Centro 2Fe-2S Centro 4Fe-4S La ferredossina Una proteina con un centro 2Fe-2S Metodo per la determinazione della sequenza dei trasportatori di elettroni In presenza di un donatore di elettroni e di O2 è stato valutato l’effetto di inibitori del trasferimento degli elettroni sullo stato di ossidazione di ogni trasportatore. In blu: composti ridotti In rosso: composti ossidati Separazione dei componenti funzionali della catena respiratoria Via seguita dagli elettroni per passare da NADH, succinato, acil-CoA e glicerolo 3-fosfato all’ubichinone (Q) NADH:ubichinone ossidoreduttasi - Complesso I (anche detta NADH deidrogenasi) NADH + Q + 5H+matrice NAD+ + QH2 + 4H+citosol Complesso II : da succinato a ubichinone Succinato Il complesso II è la succinato deidrogenasi, il solo enzima del ciclo dell’acido citrico ad essere legato alla membrana mitocondriale interna. Il complesso II contiene due tipi di gruppi prostetici e almeno 4 proteine diverse. Si pensa che gli elettroni passino dal succinato al FAD e poi attraverso i centri Fe-S all’ubichinone Il complesso di citocromi bc1 - ComplessoIII (o citocromo reduttasi) Il complesso è un dimero formato da monomeri identici, ciascuno con 11 differenti subunità. Struttura di un monomero Il centro funzionale ha tre subunità: il citocromo b (verde) con due gruppi eme (bH e bL, in rosso chiaro), la proteina ferro-zolfo di Rieske (viola) con un centro ferro-zolfo (giallo); e il citocromo c1 (blu) con un gruppo eme (rosso) Unità dimerica funzionale del Complesso III Il complesso III accoppia il trasferimento degli elettroni dall’ubichinolo (QH2) al citocromo c con il trasporto protonico dalla matrice allo spazio intermembrana. Il citocromo c1 e la proteina di Rieske sporgono verso la superficie e possono interagire con il citocromo c nello spazio intermembrana. QH2 + 2 Cyt coss + 2 H+matrice Q + 2 Cyt crid + 4H+citosol Il complesso ha due distinti siti di legame per l’ubichinone, QN e QP. Il ciclo dell’ubichinone Sul lato esterno della membrana due molecole di QH2 vengono ossidate a Q a livello del sito QP, rilasciando 4 protoni nello spazio intermembrana. Ciascuna molecola di QH2 dona un elettrone (tramite il centro Fe-S di Rieske) al citocromo c1 e un altro (tramite il citocromo b) lo dona a una molecola di Q a livello del sito QN, riducendo l’ubichinone a QH2 in due tappe successive. Questa riduzione utilizza due protoni prelevati dalla matrice. Complesso III Il funzionamento del Complesso III permette il passaggio da un trasportatore a due elettroni (l’ubichinone) a trasportatori a un solo elettrone (i citocromi b562, b566, c1 e c) e spiega la stechiometria della reazione con 4 protoni che traslocano per ciacuna coppia di elettroni che viene trasferita al citocromo c. Il risultato netto è: QH2 viene ossidato a Q e 2 molecole di citocromo c vengono ridotte. Citocromo C Il citocromo c è una proteina solubile nello spazio intermembrana. Quando il suo gruppo eme accetta un elettrone dal Complesso III, il citocromo c si sposta verso il Complesso IV per donare l’elettrone a un centro rameico binucleare di questo enzima. Subunità della citocromo ossidasi - Complesso IV La parte centrale del Complesso IV possiede tre subunità. La subunità I (giallo) ha due gruppi eme, a e a3 (rosso) e uno ione rame, CuB (sfera verde). La subunità II (blu) contiene due ioni rame (sfere verdi) complessati con i gruppi -SH di due residui di Cys in un centro binucleare, CuA, che assomiglia ai centri 2Fe-2S delle proteine ferro-zolfo. Questo centro binucleare e il sito di legame del citocromo c sono collocati in un dominio della subunità II che sporge verso lo spazio intermembrana. Non è stato ancora chiarito il ruolo della subunità III (verde) Il centro binucleare di CuA Gli ioni rame si distribuiscono equamente gli elettroni. Quando il centro è ridotto, gli ioni rame possiedono le cariche formali Cu1+Cu1+; quando è ossidato, possiedono cariche Cu1,5+Cu1,5+ . Attorno agli ioni rame sono compresi : due His (blu), due Cys (giallo), un Asp (rosso) e una Met (arancione). Il Complesso IV trasporta gli elettroni dal citocromo c all’ossigeno molecolare riducendolo ad H2O Il Complesso IV si è evoluto in modo da catalizzare una reazione redox in cui la molecola di O2 viene ridotta contemporaneamente da 4 elettroni, senza la formazione di intermedi ridotti incompleti (es.: anione superossido O2-) altamente reattivi. Flusso degli elettrononi e dei protoni attraverso i quattro complessi della catena respiratoria. Teoria chemioosmotica (Peter Mitchell, 1961) L’energia che deriva dal flusso di elettroni attraverso la catena respiratoria è utilizzata per pompare protoni (H+) dalla matrice mitocondriale verso lo spazio intermembrana. Si genera un gradiente di pH - la concentrazione di H+ è maggiore nello spazio intermembrana che nella matrice. Si genera un gradiente elettrico - la matrice è negativa rispetto allo spazio intermembrana. L’energia potenziale elettrochimica generata è chiamata forza motrice protonica. Quando i protoni fluiscono all’indietro attraverso interna tramite un enzima legato alla membrana vengono uniti a formare ATP. la membrana mitocondriale (ATP sintasi), l’ADP e il Pi Circa 3 H+ devono attraversare l’ATP sintasi per ciascuno degli ATP sintetizzati Teoria chemioosmotica Il numero di protoni pompati fuori per coppia di elettroni trasportati è 10 per il NADH e 6 per il succinato. Il numero di protoni richiesti per la sintesi di una molecola di ATP è di 4, di cui uno è utilizzato per trasportare Pi, ATP e ADP attraverso la membrana mitocondriale. Si sintetizzano 2,5 molecole di ATP (10/4) quando il donatore di elettroni è il NADH e 1,5 (6/4) quando il donatore è il succinato (FADH2). La forza motrice protonica Modello chemioosmotico F1ATPasi La subunità γ rompe la simmetria dell’esamero α3β3: ciascuna subunità β è distinta in virtù della sua interazione con una differente faccia di γ. Distinguere le tre subunità β è cruciale per il meccanismo di sintesi dell’ATP Fo contiene il canale protonico del complesso Meccanismo di sintesi dell’ATP Uno degli atomi di ossigeno dell’ADP attacca l’atomo di fosforo di Pi per formare un intermedio pentacovalente, che a sua volta forma ATP e rilascia una molecola di H2O. Diagrammi delle coordinate di reazione dell’ATP sintasi e di un tipico enzima. Nella reazione catalizzata dall’ATP sintasi la maggiore barriera energetica è il rilascio dell’ATP dall’enzima e non la sua formazione. I siti di legame per i nucleotidi della ATP sintasi non sono equivalenti La subunità γ passa attraverso il centro dell’esamero α3β3 e forma siti di legame per i nucleotidi nelle subunità β distinti l’uno dall’altro Diverse conformazioni delle subunità β Una subunità β può essere nella conformazione serrata (T), che lega l’ATP con grande avidità. Una seconda subunità β sarà nella conformazione lassa (L), che lega l’ADP e il Pi. L’ultima subunità β sarà nella conformazione aperta (O) La rotazione della subunità γ interconverte le tre subunità β favorendo la sintesi di ATP Sistemi di trasporto presenti nella membrana mitocondriale interna portano ADP e Pi nella matrice e consentono all’ATP appena sintetizzato di uscire nel citosol Il potenziale di membrana è ridotto dallo scambio ADP/ATP, che risulta nel trasferimento netto di una carica negativa fuori dalla matrice. Lo scambio ADP/ATP ha un costo energetico; circa un quarto dell’energia ricavata dal trasferimento di elettroni lungo la catena respiratoria è consumata per rigenerare il potenziale di membrana che si perde per il trasferimento dell’ATP nel citosol. Per l’ossidazione del NADH citosolico nei mitocondri, sono necessari sistemi navetta Lo shuttle del malato-aspartato Lo shuttle del glicerolo 3-fosfato Questo sistema navetta differisce da quello malato-aspartato in quanto trasporta equivalenti riducenti dal NADH (via ubichinone) al Complesso III e non al Complesso I. Per ogni coppia di elettroni che arrivano all’ossigeno dal NADH sono prodotte 2,5 molecole di ATP. Per ogni coppia di elettroni che arrivano all’ossigeno dal FADH2 sono prodotte 1,5 molecole di ATP. Le vie che producono ATP sono regolate in modo coordinato La fosforilazione ossidativa è regolata sulle necessità energetiche della cellula In condizioni fisiologiche il trasporto degli elettroni è strettamente accoppiato alla fosforilazione. Gli elettroni non scorrono attraverso la catena respiratoria se contemporaneamente non viene fosforilato l’ADP a ATP. Il fattore più importante nel determinare la velocità della fosforilazione ossidativa è la concentrazione di ADP. La regolazione della velocità della fosforilazione ossidativa (e quindi del consumo di O2) da parte dei livelli di ADP viene chiamata controllo respiratorio. Le vie che producono ATP sono regolate in modo coordinato Le concentrazioni di ATP e ADP controllano non solo le velocità di trasferimento degli elettroni e della fosforilazione ossidativa, ma anche quelle del ciclo dell’acido citrico, dell’ossidazione del piruvato e della glicolisi Generazione di calore mediante disaccoppiamento mitocondriale Il gradiente di protoni può essere cortocircuitato per produrre calore. Il tessuto adiposo bruno, mediante la proteina disaccoppiante termogenina, è specializzato in questo processo di termogenesi. I cloroplasti convertono l’energia luminosa in energia chimica Gli elettroni ad alta energia presenti nei cloroplasti vengono trasportati attraverso due fotosistemi. Durante questo tragitto, che genera potere riducente, viene sintetizzato ATP in modo analogo alla sintesi di ATP mitocondriale. Nei cloroplasti gli elettroni sono energizzati dalla luce. Rappresentazione schematica di un cloroplasto Gli eventi primari della fotosintesi avvengono nelle membrane tilacoidali La fotosintesi Il ciclo dell’acido citrico ossida a CO2 i combustibili carboniosi per generare elettroni ad alta energia. Il flusso di questi elettroni ad alta energia genera una forza motrice protonica attraverso l’azione della catena di trasporto degli elettroni. Questa forza motrice protonica viene utilizzata dalla ATP sintasi per formare ATP. Per generare elettroni ad alta energia le reazioni alla luce della fotosintesi utilizzano l’energia dei fotoni. Questi elettroni vengono utilizzati direttamente per ridurre il NADP+ a NADPH e indirettamente attraverso una catena di trasporto degli elettroni per generare una forza motrice protonica tra le due facce della membrana. Un prodotto collaterale di questa reazione è l’O2. La forza motrice protonica favorisce la sintesi di ATP attraverso una ATP sintasi, omologa a quella che interviene nella fosforilazione ossidativa Nelle reazioni al buio il NADPH e l’ATP formati dall’azione della luce favoriscono la riduzione della CO2 a composti organici più ridotti Le reazioni alla luce della fotosintesi L’energia luminosa viene assorbita e utilizzata per estrarre elettroni dall’acqua al fine di generare NADPH e per far fluire protoni attraverso la membrana tilacoidale. Questi protoni ritornano attaverso la ATP sintasi per formare ATP La clorofilla Le molecole di clorofilla (tetrapirroli con uno ione magnesio centrale) assorbono la luce in modo molto efficiente i quanto sono polieni (composti che contengono reti di legami singoli e legami doppi alternati). Un elettrone eccitato a salire in uno stato ad alta energia dall’assorbimento di un fotone può trasferirsi verso accettori di elettroni vicini. Nella fotosintesi un elettrone eccitato abbandona una coppia di molecole di clorofilla associate nota come coppia speciale. Fitolo Una molecola di fitolo, un alcol a 20 atomi di carbonio molto idrofobico, è esterificata a una catena laterale acida della clorofilla a. E’ necessario che due fotosistemi distinti assorbano fotoni perché il flusso di elettroni da acqua a NADP+ sia completo. Movimento degli elettroni e dei protoni nei tilacoidi dei cloroplasti Nel fotosistema II delle piante verdi l’eccitazione del P680, una coppia speciale di molecole di clorofilla, determina il trasferimento di elettroni al plastochinone. Gli elettroni vengono ripristinati dall’estrazione di elettroni dall’acqua in un centro contenente 4 ioni manganese. In questo centro viene generata una molecola di O2 per ogni 4 elettroni trasferiti Il plastochinolo prodotto a livello del fotosistema II viene riossidato dal complesso del citocromo bf, che trasferisce gli elettroni alla plastocianina, una rame-proteina solubile Movimento degli elettroni e dei protoni nei tilacoidi dei cloroplasti Dalla plastocianina gli elettroni entrano nel fotosistema I Nel fotosistema I l’eccitazione di una coppia speciale P700 rilascia elettroni che fluiscono alla ferredossina, un potente riducente La ferredossina -NADP+ riduttasi, una flavoproteina localizzata sul versante stromatico della membrana, converte il NADP + in NADPH. Trasferimento elettronico dall’acqua al NADP+ nella fotosintesi Questa reazione endoergonica è resa possibile dall’assorbimento della luce ad opera del fotosistema II (P680) e del fotosistema I (P700). Viene generato un gradiente protonico mentre gli elettroni attraversano il fotosistema II, il complesso bf e la ferredossina-NADP+ riduttasi. Confronto tra la fotosintesi e la fosforilazione ossidativa Nella fotosintesi il trasferimento di elettroni indotto dalla luce fa fluire protoni nel lume tilacoidale. I protoni in eccesso effluiscono da lume attraverso la ATP sintetasi per generare ATP nello stroma Nella fosforilazione ossidativa il flusso di elettroni lungo la catena di trasporto degli elettroni pompa protoni fuori dalla matrice mitocondriale. I protoni si muovono quindi dallo spazio intermembrana nella matrice attraverso la ATP sintetasi per generare ATP nella matrice. Cavolo d’Oriente Trasportatori di elettroni nella membrana mitocondriale interna delle piante
Scaricare