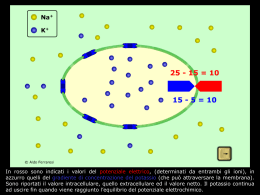
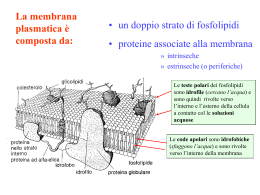
•GENESI DEL POTENZIALE DI RIPOSO DELLA MEMBRANA •POTENZIALI ELETTROTONICI E LORO PROPAGAZIONE •GENESI DEL POTENZIALE D’AZIONE •PROPAGAZIONE DEL POTENZIALE D’AZIONE IN FIBRE NERVOSE AMIELINICHE E MIELINICHE POTENZIALE DI RIPOSO DELLA MEMBRANA VR •DIFFERENZA DI POTENZIALE ELETTRICO (O VOLTAGGIO) CHE SI INSTAURA A CAVALLO DELLA MEMBRANA PLASMATICA DI OGNI CELLULA VIVENTE • VR= Vi – Ve (mV) •ASSUME VALORI NEGATIVI POTENZIALE DI RIPOSO DELLA MEMBRANA VR POTENZIALE DI RIPOSO DELLA MEMBRANA VR POTENZIALE DI RIPOSO DELLA MEMBRANA VR CAPACITA’ ELETTRICA DELLA MEMBRANA •Il doppio strato lipidico e’ in un isolante elettrico in grado di separare e accumulare ai suoi lati cariche (ioni) di segno opposto •La membrana cellulare quindi agisce come un condensatore, attuando una separazione di cariche: i cationi e gli anioni si distribuiscono ai lati opposti della membrana e interagiscono elettrostaticamente fra loro •Le soluzioni intracellulare ed extracellulare sono elettroneutre; elettroneutra è anche la regione immediatamente adiacente la membrana •La capacita’ si misura in farad (coulomb/volt); CMEMBRANA ~ 1F/cm2 DIFFUSIONE PASSIVA NETTA ATTRAVERSO UNA MEMBRANA DI IONI O MOLECOLE CARICHE •GRADIENTE CHIMICO DERIVANTE DALLE DIFFERENZE DI CONCENTRAZIONE AI DUE LATI DELLA MEMBRANA •DIFFERENZA DI POTENZIALE ELETTRICO AI CAPI DELLA MEMBRANA LA SOMMA DELLE DUE FORZE ORIGINA IL GRADIENTE ELETTROCHIMICO EQUILIBRIO ELETTROCHIMICO POTENZIALE DI EQUILIBRIO DI UNO IONE • DEFINITO COME LA DIFFERENZA DI POTENZIALE ELETTRICO TRANSMEMBRANA Ei CHE CONTROBILANCIA LA DIFFERENZA DI CONCENTRAZIONE (POTENZIALE CHIMICO) DELLO IONE i A CAVALLO DI UNA MEMBRANA PERMEABILE ALLO IONE STESSO •E’ UN POTENZIALE DI EQUILIBRIO: IL FLUSSO NETTO DELLO IONE E’ NULLO •STABILE NEL TEMPO •DIPENDE DALLA TEMPERATURA •PROPORZIONALE AL LOGARITMO DEL RAPPORTO DELLE CONCENTRAZIONI DELLO IONE AI DUE LATI DELLA MEMBRANA EQUAZIONE DI NERNST R T i2 Ei ln zi F i1 dove Ei Potenziale di equilibrio dello ione i R Costante dei gas T Temperatura assoluta zi Valenza dello ione F Costante di Faraday i Concentrazione dello ione [V] EQUAZIONE DI NERNST iout Ei 61 log iin T 310 K zi 1 [ mV] Quanto variano le concentrazioni di potassio all’equilibrio? Se Volume dei compartimenti 1 cm3 Capacità specifica della membrana 1 F cm-2 Allora avremo circa 6 x1019 ioni potassio nel compartimento 1 6 x1018 ioni potassio nel compartimento 2 Possiamo calcolare la quantità di carica Q = C V dividerla per costante di Faraday e moltiplicarla per il numero di Avogadro ottenendo circa 3,7 10 11 ioni ossia inferiore di 7 o 8 ordini di grandezza. 1 ione ogni 100 milioni si è spostato dal compartimento 1 al compartimento 2 per realizzare la condizione di equilibrio Se un canale è selettivo per uno ione i, il valore di potenziale di membrana al quale la corrente ionica è nulla (potenziale d’inversione della corrente) approssima il potenziale di equilibrio Ei Concentrazioni dei principali ioni nel muscolo scheletrico (mM) LEC LIC Potenziale di equilibrio K+ 4,5 160 -95 mV Na+ 144 7 +80 mV Ca2+ 1,3 H+ 4 10-5 (pH 7,4) 10-4 (pH 7,0) -24 mV Cl- 114 7 -80 mV HCO3- 28 10 -27 mV 10-7-10-8 M +125 - +310 mV GRADIENTE ELETTROCHIMICO Se conosciamo le concentrazioni di uno ione ai lati di una membrana e il valore di Vm possiamo calcolare il gradiente elettrochimico per lo ione i : ( Vm – Ei ) Intensità e direzione della forza che spinge lo ione ad attraversare la membrana Vr = -65 mV EK = -95 mV Vr-EK= (-65-(-95))= +30 mV Vr = -65 mV ENa = +80 mV Vr-ENa = (-65-(+80))= -145 mV IL POTENZIALE DI RIPOSO VR NON E’ UN POTENZIALE DI EQUILIBRIO E’ IL POTENZIALE AL QUALE I FLUSSI IONICI TOTALI O LA CORRENTE TOTALE CHE ATTRAVERSA LA MEMBRANA E’ NULLA EQUAZIONE DI GOLDMAN-HODGKIN-KATZ I tot 0 Per i principali ioni permeanti monovalenti: I tot I Na I K I Cl 0 RT PK K o PNa Na o PCl Cl i VR ln F PK K i PNa Na i PCl Cl o Pi Coefficiente di permeabilità della specie ionica i [cm/s] RT PK K o PNa Na o PCl Cl i VR ln F PK K i PNa Na i PCl Cl o •Le concentrazioni ioniche risultano moltiplicate per i rispettivi coefficienti di permeabilità Pi (che non figurano nell’equazione di Nernst): la differenza di concentrazione per ogni specie ionica ai due lati della membrana pesa sul valore di VR in base al valore di Pi. In altri termini VM assumerà un valore intermedio fra quello dei potenziali di equilibrio delle diverse specie ioniche avvicinandosi al potenziale di equilibrio dello ione con P maggiore •Nel caso particolare di P diversa da zero per una sola specie ionica VR coincide con il potenziale di equilibrio e l’equazione di GHK si riduce all’equazione di Nernst •V varia al variare di P GENESI DI VR Si possono distinguere processi di varia natura: •Processi di diffusione secondo gradiente elettrochimico di ioni, principalmente K+ e Na+, via canali ionici e altri trasportatori •Processo attivo che consiste nel trasporto contro gradiente elettrochimico di K+ e Na+, volto a creare e mantenere i gradienti chimici VR in quanto potenziale di diffusione tenderebbe a decadere nel tempo man mano che si dissipano i gradienti ma questo fatto non si verifica grazie all’attività della pompa Na+/ K+-ATP-asi La pompa è elettrogenica: contribuisce anche direttamente alla genesi di VR ( 3-4 mV) PROPRIETA’ ELETTRICHE PASSIVE DELLA MEMBRANA: POTENZIALI GRADUALI (ELETTROTONICI) E LORO PROPAGAZIONE Le sinapsi si formano tipicamente nei dendriti e nel soma; possono formarsi sull’assone o sul terminale assonale. 10.000 sinapsi 150.000 sinapsi •Le tracce del potenziale di membrana mostrano che sono necessari alcuni millisecondi perché si instauri e decada la polarizzazione della membrana con l’applicazione e la rimozione della corrente. •La diminuzione della velocità di salita e di caduta del potenziale indica la presenza di una capacità elettrica tra il citoplasma e il mezzo esterno. SEGNALI ELETTRICI PASSIVI NELLE MEMBRANE CELLULARI •L’INIEZIONE IN CELLULA DI UN IMPULSO DI CORRENTE RETTANGOLARE DI PICCOLA AMPIEZZA PRODUCE UNA VARIAZIONE DI VM ESPONENZIALE CHE DIPENDE DA R E DA C •LA RELAZIONE I-V ALLO STATO STAZIONARIO E’ LINEARE (LEGGE DI OHM) •SE LA MEMBRANA FOSSE RAPPRESENTATA DALLA SOLA COMPONENTE RESISTIVA ALL’INIEZIONE DI UN IMPULSO DI CORRENTE RETTANGOLARE DOVREBBE CORRISPONDERE UNA VM RETTANGOLARE E ISTANTANEA •VM NON E’ ISTANTANEA PERCHE’ RALLENTATA DALLA CAPACITA’ DELLA MEMBRANA VM (t ) I m R(1 e dove R C costante di tempo t / ) SEGNALI ELETTRICI GENERATI NEI NEURONI POTENZIALI GRADUALI •SONO SEGNALI LOCALI: PERCORRONO BREVI DISTANZE DAL PUNTO DI ORIGINE •SONO DEPOLARIZZAZIONI O IPERPOLARIZZAZIONI •AMPIEZZA PROPORZIONALE ALL’INTENSITA’ DELLO STIMOLO CHE LI GENERA •SI VERIFICANO TIPICAMENTE NEI DENDRITI E NEL SOMA DI UN NEURONE •NEL SNC E IN NEURONI EFFERENTI SI GENERANO QUANDO SEGNALI CHIMICI PROVENIENTI DA ALTRI NEURONI APRONO CANALI IONICI REGOLATI CHIMICAMENTE •SI PROPAGANO DIREZIONI CON DECREMENTO LUNGO TUTTE LE PROPAGAZIONE ELETTROTONICA • IL POTENZIALE GRADUALE (ELETTROTONICO) SI PROPAGA LUNGO TUTTE LE DIREZIONI MA LA SUA AMPIEZZA DIMINUISCE ESPONENZIALMENTE IN FUNZIONE DELLA DISTANZA PERCORSA DAL PUNTO DI INIEZIONE DELLA CORRENTE CHE LO HA GENERATO a) Dispersione della corrente attraverso la membrana plasmatica dovuta alla presenza di canali ionici aperti che lasciano uscire cariche positive nel mezzo extracellulare b) Resistenza del citoplasma al flusso di corrente Vx V0e V0 x/ Variazione del potenziale nel punto x=0 Vx Variazione del potenziale alla distanza x Costante di spazio Rm Rl R m Resistenza di un’unità di lunghezza della membrana assonale Rl Somma delle resistenze longitudinali interna ed esterna a a = raggio dell’assone SEGNALI ELETTRICI GENERATI NEI NEURONI POTENZIALI GRADUALI O POTENZIALI LENTI •SONO INTEGRATI NELLA ZONA TRIGGER NEURONE DOVE POSSONO GENERARE POTENZIALE D’AZIONE DEL UN •SONO POTENZIALI GRADUALI I POTENZIALI POSTSINAPTICI ECCITATORI (EPSP) E I POTENZIALI POSTSINAPTICI INIBITORI (IPSP) SEGNALI ELETTRICI GENERATI NEI NEURONI POTENZIALI D’AZIONE •SEGNALI ELETTRICI RAPIDI E TRANSITORI CHE PROPAGANO VELOCEMENTE SENZA PERDERE INTENSITA’ SI •SONO DEPOLARIZZAZIONI DI BREVE DURATA DI CIRCA 100 mV DI AMPIEZZA •SI VERIFICANO NELLA ZONA TRIGGER DEL NEURONE IN RISPOSTA AD UN VALORE CRITICO DI DEPOLARIZZAZIONE DEL POTENZIALE DI MEMBRANA DETTO VALORE SOGLIA •SONO FENOMENI “TUTTO O NULLA” CANALI IONICI GENESI DEL POTENZIALE D’AZIONE RIPOLARIZZATO RIPOLARIZZATO 1. POTENZIALE DI MEMBRANA A RIPOSO 2. INIZIO DELLA FASE DI DEPOLARIZZAZIONE (VOC DEL Na+ e K+ SI APRONO) 3. SOGLIA 4. CICLO DI HODGKIN A RETROAZIONE POSITIVA: TUTTI I CANALI DEL Na SI APRONO 5. INATTIVAZIONE DI VOC DEL Na + 6. MASSIMA ATTIVAZIONE DI VOC DEL K + 7. VOC DEL K+ ANCORA APERTI 8. VOC DEL K+ SI CHIUDONO E Na + ENTRA IN CELLULA 9. SI RISTABILISCE VR CICLO DI HODGKIN PERIODO REFRATTARIO ASSOLUTO E RELATIVO PROPAGAZIONE DEL POTENZIALE D’AZIONE PROPAGAZIONE SALTATORIA STRUTTURA E FUNZIONE DEI CANALI IONICI VOLTAGGIO DIPENDENTI Filtro di selettività situato lungo la parete interna del canale: caratterizzato dalla presenza di un sito specifico che lega blandamente gli ioni sodio. Secondo questo modello, uno ione sodio, quando attraversa il poro, si lega reversibilmente ad un sito attivo. La carica positiva dello ione viene stabilizzata nel sito di legame dal residuo idrofilo di un aminoacido caricato negativamente che costituisce una delle pareti del canale e da una molecola di acqua che viene attratta dal residuo di un secondo aminoacido posto sulla parete opposta del poro. Secondo il modello, gli ioni potassio che hanno dimensioni maggiori, non sono in grado di attraversare la zona del filtro, in quanto le interazioni elettrostatiche con le cariche negative del filtro stesso non li stabilizzano in maniera efficace L’apertura dei canali voltaggio-dipendenti comporta una ridistribuzione di cariche all’interno del segmento S4. Modello della regione S4 per il canale del sodio: gli R rappresentano i residui di arginina carichi positivamente. A riposo ogni carica positiva netta della struttura ad -elica è stabilizzata da una carica negativa presente nelle regioni vicine della molecola. Quando la cellula viene depolarizzata, la variazione dela campo elettrico determina uno spostamento delle cariche positive verso l’estremità esterna della membrana (gating currents), con un andamento elicoidale a passo di vite. Il movimento si arresta quando le cariche in S4 incontrano altre cariche negative fisse di strutture elicoidali adiacenti. Il riallineamento delle cariche stabilizza la nuova conformazione del canale. Canale del sodio voltaggio-dipendente Subunità Canale del calcio voltaggio-dipendente Subunità Canale del potassio voltaggio-dipendente Subunità •Almeno 40 membri divisi in 12 sottofamiglie
Scaricare




![e /[K]](http://s2.diazilla.com/store/data/000059268_1-35d765178255b5408638f1bb10cad1ef-260x520.png)