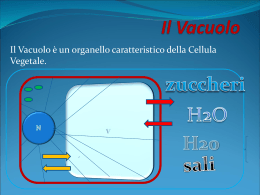
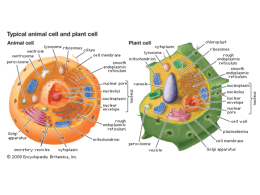
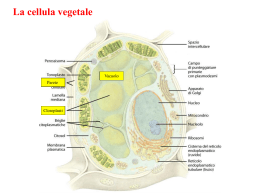
METABOLISMO CELLULARE METABOLISMO PRIMARIO: tutti quei processi che sono fondamentali per la vita dell’organismo sia vegetale che animale e quindi: sintesi di macromolecole (quali es. proteine, acidi nucleici, lipidi e zuccheri), respirazione, fotosintesi, traspirazione trasporto METABOLISMO SECONDARIO: tutti quei processi NON ESSENZIALI per la vita dell’organismo. Si definiscono metaboliti secondari o prodotti secondari quei composti presenti negli organismi vegetali che in genere rappresentano prodotti di vie metaboliche che apparentemente non hanno un ruolo di base (ma secondario) nel metabolismo della cellula. Molti di questi composti sono tuttavia ecologicamente importanti nell’interazione tra la pianta e l’ambiente o tra la pianta ed altri organismi con i quali è in competizione. VACUOLO Il VACUOLO è presente SOLO nelle cellule vegetali. Mentre le cellule vanno assumendo la forma e le dimensioni definitive, compaiono nel citoplasma numerosi piccoli VACUOLI, che aumentano sempre di volume, durante la crescita delle cellule vegetali. Il vacuolo è rivestito da una membrana detta TONOPLASTO e contiene una soluzione acquosa detta SUCCO VACUOLARE TONOPLASTO Il TONOPLASTO è una membrana lipoproteica bistratificata ASIMMETRICA: la superficie rivolta verso l’esterno è ricca di proteine intramembrana rispetto alla faccia rivolta verso l’interno. Le proteine intramembrana sono per la maggior parte proteine carriers, pompe protoniche, proteine canali ed enzimi. Spesso di tratta di GLICOPROTEINE cioè proteine complessate con residui oligosaccaridici rivolti verso il succo vacuolare. Inoltre il DOPPIO STRATO LIPIDICO del tonoplasto presenta in minima parte fosfolipidi mentre sono predominanti i GLICOLIPIDI (es. galattolipidi). Tra le pompe protoniche molto importanti sono le H+ATPasi e H+-PPasi le quali idrolizzano adenosina trifosfato (ATP) e pirofosfato inorganico (PPi) generando una forza protonica per il trasporto di ioni e metaboliti attraverso il tonoplasto. GENESI e DIFFERENZIAMENTO DEL VACUOLO Le cellule non differenziate normalmente posseggono iniziali vacuolari dette PROVACUOLI; il processo di formazione del vacuolo normalmente inizia durante le primissime fasi del differenziamento. Si ritiene che i vacuoli abbiano origine da una porzione del reticolo endoplasmatico, localizzata in prossimità della faccia trans dei sacculi del Golgi, costituita da un sistema tubulare membranoso interconnesso a cisterne appiattite. Faccia trans dell’Apparato del Golgi Questa particolare porzione di reticolo endoplasmatico è stata identificata con la sigla di GERL (G = Golgi, ER = Reticolo Endoplamstico; L = Lisosoma). Come si formano i vacuoli? Ci sono diverse teorie; partecipano porzioni dell'apparato di Golgi, reticolo endoplasmatico liscio e i lisosomi (corpiccioli delimitati da una membrana che contengono vescicole di enzimi latici). I 3 si dispongono su una superficie di una sfera, intorno c'è citoplasma e organelli. Tubuli, le cisterne e le vescicole si fondono costituendo 2 membrane: quella esterna è resistente, quella interna viene digerita così da formare un'unica membrana: il tonoplasto contenente del succo cellulare; in tal modo possono costituirsi tanti vacuoli. Il GERL di norma si espande in tubuli allungati e ramificati in cui si accumulano enzimi idrolitici. Questi tubuli sono i provacuoli nascenti e visto che contengono idrolasi acide possono considerarsi quasi come lisosomi (generalmente formati dal Golgi). I provacuoli intraprendono fenomeni d’autofagia: si allungano in strutture tubulari e ramificate che progressivamente si avvicinano e fondendosi delimitano in maniera continua, attraverso un doppio strato di membrana, una porzione di citoplasma (I, II) l’azione di enzimi idrolitici determina l’idrolisi sia del citoplasma che della membrana interna, si ha così la formazione di una struttura sferica (VACUOLO AUTOFAGICO) delimitata da una membrana detta tonoplasto (III) con il differenziamento cellulare, i piccoli vacuoli si ingrandiscono sia attraverso la fusione di vescicole prodotte dall’apparato del Golgi e del reticolo endoplasmatico, sia fondendosi tra loro per formare il grande vacuolo della cellula differenziata (IV, V) FUNZIONI DEI VACUOLI I vacuoli svolgono importantissime funzioni: 1) RUOLO OSMOTICO (insieme al citoplasma ed alla parete cellulare) a) SUPPORTO MECCANICO: il vacuolo insieme alla parete realizza una struttura rigida (la pressione dell’acqua nel vacuolo viene controbilanciata dalla rigidità della parete cellulare) che determina la PRESSIONE DI TURGORE responsabile sia della distensione cellulare sia della rigidità di tessuti non lignificati (es. foglie, giovani fusti) i quali quando la pressione di turgore diminuisce avvizziscono. Infatti se la parete non fosse in grado di opporsi al continuo richiamo d’acqua si avrebbe lo scoppio dei protoplasti e se il vacuolo perdesse acqua, determinando l’abbassamento della pressione di turgore, si avrebbe la PLASMOLISI cellulare. b) FORZA MOTRICE PER LA DISTENSIONE CELLULARE La pressione di turgore esercitata dal vacuolo in maniera uniforme sulla parete cellulare rappresenta la forza guida dell’accrescimento delle cellule vegetali Nei vegetali l’uscita da questo ciclo, l’abbandono dello stato meristematico e l’inizio della differenziazione comporta sempre l’effettuazione di un ulteriore tipo di crescita, l’accrescimento per distensione, che modifica profondamente dimensioni, forma e struttura delle cellule. Le dimensioni, prima di tutto : se una cellula meristematica ha un diametro di 15-20µm una cellula adulta che si rispetti è grande almeno 150-200µm (ma alcune cellule possono essere lunghe mm)(Fig.21) La forma : dall’aspetto poliedrico, più o meno isodiametrico, si passa a forme in genere allungate, con una dimensione decisamente superiore alle altre . La struttura interna: se durante la crescita embrionale , nella cellula che raddoppia di volume, si assiste a una sintesi di tutti i componenti cellulari , nella crescita per distensione il grosso dell’aumento di volume (che può essere di centinaia di volte ) è dovuto alla parete cellulare e al vacuolo, con un contributo meno rilevante dei plastidi. Poiché il vacuolo è costituito principalmente di acqua, possiamo dire che la crescita per distensione è caratterizzata da un massiccio ingresso di acqua nella cellula . Come già sapete l’assunzione e la perdita di acqua da parte di una cellula avvengono per osmosi. Lo stato di turgore di una cellula è la risultante di una condizione di equilibrio fra il potenziale osmotico della cellula (dovuto ai soluti disciolti) e il potenziale di pressione, dovuta alla parete, elastica si ma non estensibile in modo plastico irreversibile. Nella condizione di equilibrio fra i due potenziali , ogni ulteriore ingresso di acqua è impossibile, a meno che la parete non venga indebolita tanto da consentire alle cellule di estenderla in modo irreversibile, facendone aumentare la superficie e diminuire il potenziale di pressione. DIVISIONE CELLULARE DISTENSIONE CELLULARE 2) RUOLO di RISERVA di ACQUA: Fenomeno della PLASMOLISI Ambiente ISOTONICO = concentrazione dei soluti interna uguale a quella esterna Ambiente IPOTONICO = concentrazione dei soluti interna maggiore di quella esterna (entra in gioco la parete cellulare e si ha TURGORE CELLULARE) Ambiente IPERTONICO = concentrazione dei soluti interna minore a quella esterna (fenomeno della PLASMOLISI) Soluzione ISOTONICA Soluzione IPERTONICA 3) RUOLO TAMPONANTE del pH citoplasmatico. Il succo vacuolare ha un pH acido il cui valore in condizioni fisiologiche normali si aggira tra pH 4 e 5. Questo valore è sempre notevolmente più basso rispetto a quello del citoplasma (pH 6.8-6.9). La capacità di mantenere il succo vacuolare ad un pH acido è dovuto alla presenza sul tonoplasto di pompe protoniche (H+-ATPasi e H+-PPasi) che attraverso l’idrolisi dell’ATP e del pirofosfato inorganico forniscono protoni al succo vacuolare acidificandolo. La presenza di pompe protoniche sul tonoplasto è di notevole importanza fisiologica poiché alla loro funzionalità è strettamente legata la capacità di trasportare selettivamente ioni e metaboliti dal citoplasma al vacuolo e, in misura minima dal vacuolo al citoplasma, permettendo così il mantenimento del pH costante del citoplasma. 4) EQUILIBRIO e RISERVA DI IONI Nel vacuolo si accumulano molti ioni inorganici la cui natura e quantità è dovuta al tipo di terreno su cui cresce la pianta. Tutti gli ioni non immediatamente utilizzati vengono immagazzinati nel vacuolo mediante l’utilizzo di speciali proteine trasportatrici a livello del TONOPLASTO (TRASPORTO ATTIVO). Piante che vivono su terreni ricchi di elementi tossici tendono ad accumularli nel vacuolo. Gli ioni nel vacuolo possono rimanere sotto forma libera oppure formare dei sali e dei cristalli con gli acidi organici pure accumulati nel vacuolo (es. ossalato di Calcio: acido ossalico + Ca2+). In particolare cristalli di ossalato di calcio formano degli inclusi solidi insolubili di varie forme (rafidi, prismi o stiloidi, druse, sabbia cristallina). 5) RISERVA di METABOLITI Molti metaboliti organici di riserva vengono accumulati nel vacuolo tra cui aminoacidi e proteine, zuccheri monosaccaridi (es. glucosio nell’uva), disaccaridi (saccarosio nella barbabietola e nella canna da zucchero), pigmenti che conferiscono i colori ai fiori alla frutta etc. (flavonoidi, antociani, flavoni e a volte tannini). Gli acidi organici vengono accumulati nel vacuolo per rimuoverli da citoplasma quando sono in eccesso in quanto possono interferire in varie vie metaboliche es. acido citrico (agrumi), acido malico, acido ossalico. Nelle cellule dei tessuti di riserva dei semi accumulare formare i GRANULI DI ALEURONE che sono contenuti in vacuoli fortemente modificati per immagazzinare proteine che vengono convogliate probabilmente attraverso il reticolo endoplasmatico, e che precipitano nel vacuolo durante la maturazione a causa della disidratazione. I granuli d’aleurone possono contenere, oltre alle sostanze proteiche, anche altre sostanze come ad es. la fitina. Possiedono una membrana che delimita una MATRICE PROTEICA (formata da albumine) che può includere un CRISTALLOIDE PROTEICO (formato da globuline), e corpi sferoidali di fitina (GLOBOIDI). Durante la germinazione le riserve vengono mobilizzate e i granuli aleuronici si dilatano per formare vacuoli pressoché normali. 6) SEGREGAZIONE DI METABOLITI Alcuni METABOLITI SECONDARI vengono riversati nel vacuolo come sostanze di rifiuto anche se vengono poi riutilizzati in altre vie metaboliche. Queste sostanze sono: a) ALCALOIDI: foltissimo gruppo di sostanze organiche contenenti azoto di norma in anelli eterociclici, in genere a reazione basica ed inodori. In genere sono veleni o importanti principi attivi di piante medicinali di piante medicinali e possono avere importanti effetti biologici su piante, parassiti ed animali. Esempi colchicina e vinblastina bloccano la proliferazione cellulare; atropina, scopolamina, caffeina, papaverina, cocaina, efedrina, morfina, nicotina agiscono a livello del sistema nervoso centrale e periferico; stricnina, aconitina hanno effetti tossici. b) GLICOSIDI: sono composti formati da uno zucchero (il più comune è il glucosio) complessato ad una molecola non zuccherina. In generale sono sostanze di sapore amaro e abbastanza diffuse. Possono essere anche molto tossici es. glicosidi cianogenici che liberano per idrolisi acido cianidrico. I glicosidi sono presenti nelle mandorle amare, mele, pesche, susine (amigdalina), nel sambuco, nel granoturco e nel sorgo (murrina), in varie Leguminose. c) TERPENI: sono lipidi (quelli a basso peso melocolare sono volatili), profumati, insolubili in acqua. I terpeni sono i principali componenti degli OLI ESSENZIALI e conferiscono alle piante un profumo ed un’aroma spesso gradevole. Gli oli essenziali sono presenti nei fiori, nelle foglie, nelle foglie e nei frutti es. Labiate cioè piante aromatiche (basilico, salvia etc), finocchio, sedano, prezzemolo. Le PIRETRINE sono un’altra classe di terpeni presenti nelle foglie e nei fiori es di crisantemo, che esercitano una forte tossicità verso certi insetti (piretro negli insetticidi). d) Alcuni vacuoli contengono: RESINE, OLEORESINE (trementine), BALSAMI, GOMMORESINE (es. mirra e incenso), GOMME NATURALI (es. caucciù dall’Hevea brasiliensis che è un politerpene). 7) FUNZIONI LITICHE La presenza di idrolasi acide (peptidasi, glicosidasi, esterasi) dentro il vacuolo fa sì che esso venga considerato il più importante compartimento litico della cellula vegetale. Quando i vacuoli hanno funzione idrolitica sono analoghi ai LISOSOMI delle cellule animali. Quando gli organuli cellulari invecchiano o si deteriorano si fondono con il tonoplasto e sono trasferiti nel vacuolo dove vengono demoliti dagli enzimi idrolitici (portati da vescicole del Golgi) presenti all’interno. Le sostanze liberate (zuccheri, aminoacidi, nucleotidi, etc.) vengono presumibilmente trasportate al di fuori del vacuolo e riutilizzate. Consente alla cellula di raggiungere notevoli dimensioni Evita la formazione di spazi vuoti Spinge il citoplasma verso l'esterno della cellula facilitando gli scambi metabolici Rappresenta un sistema di escrezione dei rifiuti Regola l'omeostasi, funzionando come osmometro, mediante variazioni di concentrazione del succo vacuolare Funziona da organulo di riserva (acqua e varie sostanze) Concorre alla colorazione di fiori, frutti e altre parti vegetali 1)Ioni minerali Na +, K+, Ca ++, Mg ++, Cl-, SO?H?PO?-, NO?-. Sono importanti per la pressione osmotica. 2) Prodotti del metabolismo primario perché comuni a tutte le piante Glucosio (uva, ciliegie) Fruttosio (mele, pere) Saccarosio (canna da zucchero, barbabietola) Inulina (composite) 3) Acidi organici: acido malico, acido assolico, acido citrico, acido ascorbico . 4) Prodotti del metabolismo secondario Tannini: sostanze scure nella corteccia di molte piante, molto complessi, hanno compito deterrente; l'uomo li usa per conciare le pelli Pigmenti: antociani, flavoni, il colore dipende dal PH della soluzione vacuolare Terpeni: complesse sostanze volatili profumate, servono per richiamare gli insetti; impollinazione nell'industria cosmetica Glicosidi: amigdalina, vicianina, digitalina (molecole complesse: una parte è costituita da uno zucchero, l'altra è varia. Spesso la parte che lega zucchero ha attaccato un radicale cianidrico: glicosidi cianogenetici : sono confinati nel vacuolo mentre nel citoplasma ci sono enzimi in grado di liberare il cianuro :leguminose) Alcaloidi: nicotina, atropina, caffeina, cocaina, morfina (molecole complesse che contengono azoto) Inclusi vacuolari solidi: - ossalato di calcio: sono cristalli duri (druse) - aleurone. Proteine di riserva accumulate nei semi soprattutto dalle leguminose
Scaricare