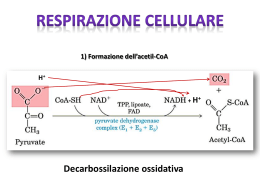
RESPIRAZIONE CELLULARE ATP VIE GLICOLITICHE PDH CH3COCOOH CATENA RESPIRATORIA NAD+/FAD CH3COSCoA CICLO DI KREBS CO2 + NADH/FADH2 Destino del piruvato……. Il piruvato entra nel mitocondrio grazie ad una proteina trasportatrice,per essere decarbossilato ad Acetil CoA ed entrare nel ciclo di krebs Destino aerobico del Piruvato L’acido piruvico o il lattato vengono completamente ossidati a CO2 attraverso il ciclo dell’acido citrico (TCA cycle), con conseguente rilascio di energia sotto forma di ATP e di trasportatori di elettroni ridotti, principalmente NADH È utile pensare all’ossidazione metabolica dei substrati come a un processo a 3 stadi: • il carbonio è incorporato nell’acetil-CoA • il carbonio viene quindi ossidato a CO2, trasportatori ridotti di elettroni e una piccola quantità di ATP • i trasportatori ridotti di elettroni sono riossidati fornendo energia per la sintesi di ulteriore ATP (fosforilazione ossidativa) L’attività del ciclo TCA è favorita da bassi rapporti NADH/NAD+ Il ciclo degli acidi tricarbossilici e’ la via finale comune per l’ossidazione delle molecole combustibili (glucosio, amminoacidi, acidi grassi). Il ciclo dell’acido citrico include una serie di ossidoriduzioni che portano all’ossidazione di un gruppo acetile a due molecole di CO2. PIRUVATO La funzione del ciclo di Krebs è quella di immagazzinare elettroni ad alta energia provenienti da combustibile organico. CO2 Acetil CoA Acetil CoA+ 3 NAD+ + FAD + GDP + Pi +2H2O 2CO2+ GTP + 3NADH +3H+ + CoA + FADH2 GTP Ciclo di Krebs 8 e- CO2 Il Ciclo di Krebs avviene nella matrice mitocondriale 1° STEP: Formazione del citrato Condensazione di una unità C4 (ossalacetato) con una unità C2 (gruppo acetile dell’acetil CoA) enzima: citrato sintasi L’equilibrio di questa reazione è tutto spostato verso la formazione dell’acido citrico dalla elevata energia che si libera dal legame tioestereo dell’acetil-CoA. Quando la carica energetica cellulare aumenta, il citrato tende ad accumularsi. L'eccesso di citrato viene usato per trasportare all'esterno del mitocondrio l'acetil-CoA. CITRATO SINTASI MECCANISMO D’AZIONE Il meccanismo prevede un attacco nucleofilo dell’enolato del tioestere al gruppo carbonilico dell’ossalacetato con formazione di citrato CITRATO SINTASI MECCANISMO D’AZIONE Nel sito catalitico (due per enzima) è presente un residuo di aspartato che, insieme ad un residuo di istidina, contribuisce a mantenere in sede la molecola di acetil-CoA attraverso interazioni ioniche. Tali residui contribuiscono ad attivare la molecola ad un intermedio enolico, rendendola affine per un centro carbonioso dell'ossalacetato Idrolisi leg.tioestere La sua struttura quaternaria consta di due subunità, ad ognuna delle quali si possono legare i due substrati. Il sito attivo dell'enzima attiva l'acetil-CoA per renderlo affine ad un centro carbonioso dell'ossalacetato. In seguito al legame tra le due molecole, il gruppo tioestere (CoA) viene idrolizzato, formando così la molecola di citrato. La reazione è altamente esoergonica (ΔG'°=-30 kJ/mole), motivo per cui questo step è irreversibile. Il citrato prodotto dall'enzima, inoltre, è in grado di inibire competitivamente l'attività dell'enzima. Pur essendo la reazione molto favorita (perché esoergonica), dunque, la citrato sintasi può essere saldamente regolata. Questo aspetto ha una notevole importanza biologica, dal momento che permette una completa regolazione dell'intero ciclo di Krebs, rendendo l'enzima una sorta di pacemaker dell'intero ciclo. La citrato sintetasi è sede di regolazione allosterica. NADH Succinil CoA - EFFETTORI allosterici negativi CITRATO Il citrato può alimentare le successive tappe del ciclo di Krebs oppure uscire dal mitocondrio nel citoplasma, dove in una reazione inversa a quella della condensazione produce l’acetil-CoA necessario per la sintesi degli acidi grassi. Il citrato può funzionare anche come: TRASPORTATORE DELL’ACETILE attraverso la membrana mitocondriale; EFFETTORE NEGATIVO della fosfofruttochinasi, inibendo la glicolisi; EFFETTORE POSITIVO della Acetil-CoA carbossilasi stimolando la biosintesi degli acidi grassi. 2° STEP: Isomerizzazione del citrato in isocitrato Enzima: aconitasi L'enzima catalizza anche la reazione inversa, ma nel ciclo di Krebs tale reazione è unidirezionale a causa della legge di azione di massa: le concentrazioni (in condizioni standard) di citrato (91%), dell'intermedio cis-aconitato (3%) e di isocitrato (6%), infatti, spingono la reazione decisamente verso la produzione di isocitrato. ACONITASI - Meccanismo di azione E’ un'eliminazione di H20 favorita dalla formazione di un acido α,β insaturo in equilibrio con l’idratazione ad acido isocitrico 3° STEP:Decarbossilazione ossidativa dell’isocitrato Ossidazione decarbossilazione enzima: isocitrato deidrogenasi Il processo consta di due fasi:la prima coinvolge l'ossidazione dell'isocitrato (un alcool secondario) in ossalsuccinato (un chetone), seguita dalla decarbossilazione del gruppo carbonile b nel chetone, formando a-chetoglutarato. ISOCITRATO DEIDROGENASI Un'altra isoforma dell'enzima catalizza la stessa reazione, non nel ciclo di Krebs ma nel citosol e nei mitocondri, ed utilizza come cofattore il NADP+ al posto del NAD La struttura dell’enzima non è stata ancora ben definita. Attualmente si ipotizza che la proteina sia composta da 3 subunità, sia allostericamente regolata, e che richieda come sustrato ione Mg2+ o Mn2+. L'omologo più simile che abbia una struttura conosciuta è l' E. coli NADP-dipendente Isocitrato deidrogenasi (IDH), che ha solamente 2 subunità e un 13% di identità e 29% di somiglianza basata sulle sequenze di amminoacidi, facendone un lontano parente del IDH umano e non attendibile per una comparazione certa. . ISOCITRATO DEIDROGENASI (meccanismo di azione) Lo step dell'IDH, a motivo del suo grande dispendio di energia liberata, è una delle reazioni irreversibili nel ciclo di Krebs e per questo deve essere accuratamente regolata per evitare un'inutile dispersione di isocitrato (e un conseguente accumulo di α-chetoglutarato). REGOLAZIONE ALLOSTERICA isocitrato, NAD+, Mg2+ / Ca2+, ADP + NADH e a-chetoglutarato, ATP - NADH e ATP Forma attiva NAD+ e ADP Forma inattiva 4° STEP:Decarbossilazione ossidativa dell’a-chetoglutarato Enzima: a-chetoglutarato deidrogenasi La decarbossilazione ossidativa dell’ a-chetoglutarato in succinil- CoA è identica alla decarbossilazione del piruvato in Acetil-CoA come meccanismo. Il complesso enzimatico a-chetoglutarato deidrogenasi è associato analogamente agli stessi coenzimi: TPP, acido lipoico,FAD, NAD+ e CoA. A differenza della piruvato DH non è regolata dal processo “fosforilazionedefosforilazione”. La reazione è irreversibile. Formazione di una seconda molecola di CO2 Tale reazione (decarbossilazione ossidativa a succinil –CoA) mostra la liberazione di una molecola di CO2, espressione della completa degradazione ossidativa dell’acetile immesso nel ciclo. L’a- chetoglutarato, non rappresenta solo un intermedio del ciclo di Krebs ma è anche metabolita importante di confluenza del METABOLISMO GLUCIDICO E DEGLI AMINOACIDI. Può essere infatti trasformato in glutammato, così come il glutammato può essere convertito in a- chetoglutarato dalle transaminasi. 5° STEP:Trasformazione del Succinil-CoA in Succinato fosforilazione a livello di substrato Enzima: succinil-CoA sintetasi o succinato tiochinasi L’energia del legame tioestereo che si viene a rompere è trasferita nel legame pirofosforico del neoformato GTP. Questa è l’unica reazione metabolica ove il GTP è preferito all’ATP quale accettore primario di un gruppo fosforico ricco di energia. 6° STEP:Ossidazione del succinato succinato deidrogenasi La succinato deidrogenasi (o succinato-coenzima Q riduttasi) è un enzima complesso che si trova nella matrice della membrana mitocondriale interna. È un enzima chiave del ciclo di Krebs ed è l'unico di questo ciclo ad essere legato ad una membrana, cio’ consente il trasferimento diretto degli elettroni alla catena respiratoria, sita nella membrana. SUCCINATO DEIDROGENASI La succinato deidrogenasi è un eterotetramero diviso in 3 domini: SdhA, il dominio catalitico; SdhB, la subunità per il trasferimento di elettroni; SdhCD, che contiene eme di tipo b, la cui funzione non è ancora chiara. Il dominio SdhA contiene il cofattore FAD, legato covalentemente ad un residuo conservato di istidina, che è il sito in cui dove il succinato viene ossidato in fumarato. Questo causa la riduzione del FAD in FADH2. La subunità per il trasferimento di elettroni (SdhB) contiene un cluster [2Fe-4S], uno [4Fe-4S] ed uno [3Fe-4S], che favoriscono il passaggio di elettroni dall'SdhA al dominio di membrana. Questi elettroni vengono indirizzati, per la riduzione di una molecola di ubiquinone su un sito quinone-legante collocato nel dimero SdhCD. La riduzione del quinone è un processo a due elettroni e richiede la formazione di un intermedio, il radicale ubisemiquinone. La risultante molecola di ubiquinolo viene rilasciata, libera di diffondersi attraverso la membrana cellulare e di interagire con i successivi enzimi della respirazione cellulare 7°STEP: idratazione del fumarato Il deficit di fumarasi, detto anche aciduria fumarica, è una malattia genetica rarissima, autosomica recessiva dovuta a mutazioni inattivanti sul gene della fumarasi. La malattia è estremamente rara: ne sono stati rilevati solo 33 casi nel mondo di cui addirittura 20 negli ultimi anni in due comunità della Chiesa di Gesù Cristo dei Santi degli Ultimi Giorni (nelle periferie di Colorado City e Hildale) il cui fondatore comune, Joseph Smith Jesop, o sua moglie, erano portatori sani del gene mutato. A causa del carattere chiuso di tali comunità e delle pratiche di poligamia e matrimoni tra consanguinei, si è verificata un'anormale moltiplicazione della sequenza genica difettata e quindi un aumento innaturale della frequenza dell'omozigosi, ovvero della malattia. Malattie legate alla succinato deidrogenasi Le malattie legate alla funzionalità della succinato deidrogenasi includono il paraganglioma ereditario, disordini ossessivo-compulsivi, emissioni di flatulenze aromatiche e nanismo. La rimozione di questo enzima dal genoma degli embrioni si è dimostrata letale. 8°STEP:Ossidazione del malato Ciclo di krebs RESA NETTA IN ATP DAL METABOLISMO AEROBIO DEL GLUCOSIO 6 6 18 4 24 ATP Il ciclo di Krebs è sia un processo catabolico che anabolico Alcune vie biosintetiche utilizzano intermedi del ciclo Gli intermedi sottratti devono essere rimpiazzati REGOLAZIONE DEL CICLO DI KREBS Le tre reazioni chiave del ciclo sono quelle fortemente esoergoniche ENZIMA DG°' DG (kJ/mol) (kJ/mol) CITRATO SINTASI - 31,5 negativo ACONITASI circa 5 circa 0 ISOCITRATO DEIDROGENASI - 21 negativo a-CHETOGLUTARATO DEIDROGENASI - 33 negativo SUCCINIL - CoA SINTETASI - 2,1 circa 0 SUCCINATO DEIDROGENASI +6 circa 0 FUMARASI -3,4 circa 0 + 29,7 circa 0 MALATO DEIDROGENASI Questo schema del ciclo dell’acido citrico indica i punti di inibizione (indicati con il circolo con il segno-) e gli intermedi della via che agiscono da inibitori (linee rosse). Ca++ e ADP sono invece attivatori La citrato sintasi è inibita da citrato, NADH e succinil-CoA L'isocitrato deidrogenasi è inibita da NADH e ATP a-chetoglutarato deidrogenasi è inibita da NADH e succinilCoA Calcio ed ADP attivano l'isocitrato deidrogenasi Calcio attiva deidrogenasi sia la PDH che l'a-chetoglutarato REGOLAZIONE DEL CICLO DI KREBS Citrato sintetasi - Citrato, succinil CoA, ATP,NADH INIBIZIONE Isocitrato deidrogenasi - ATP, NADH - ADP, NAD+ Ca++ INIBIZIONE ATTIVAZIONE a- chetoglutarato - NADH, Succinil CoA - Ca++ INIBIZIONE ATTIVAZIONE REAZIONI ANAPLEROTICHE : Reazioni che apportano metaboliti essenziali al ciclo. 1. Alcuni aminoacidi------> alfa-Chetoglutarato. 2. Acidi grassi a catena dispari ------> SuccinilCoA 3. Isoleucina,Metionina,Valina ------> SuccinilCoA 4. Tirosina, Fenilalanina, Aspartato ------> Fumarato 5. Alcuni aminoacidi ------> Ossaloacetato 6. Piruvato ------> Ossaloacetato in celeste a.a. corpi chetonici in giallo a.a glucosio in rosa a.a. glucosio e corpi chetonici arginina, glutammina, istidina, prolina glicina, alanina, serina, cisteina,triptofano isoleucina glutammato piruvato propionil~CoA isocitrato triptofano leucina acetil-CoA isoleucina citrato acetoacetil-CoA ossalacetato leucina lisina fenilalanina tirosina malato aspartato, asparagina a-chetoglutato biotina B12 succinil~CoA succinato fumarato fenilalaninatir osina valina metionina treonina
Scaricare