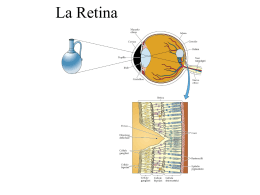
Fotorecezione Dr. Paola Perin Dipartimento di Scienze Fisiologiche-Farmacologiche Cellulari-Molecolari – Università di Pavia Cosa e’ la luce • Luce visibile: radiazioni elettromagnetiche con l tra 400 e 700 nm • I raggi luminosi viaggiano in linea retta nel vuoto o in un mezzo isotropo • Alla superficie di contatto tra due mezzi si puo’ avere: – Rifrazione (mezzi trasparenti) – Riflessione (mezzi opachi) – Assorbimento (dipende dalla struttura chimicofisica) Ma noi cosa vediamo? Luce riflessa o prodotta dall’oggetto Intensita’ di illuminazione Quantita’ di luce che entra nell’occhio • La visibilita’ di un oggetto dipende dall’intensita’ dei raggi luminosi che ne escono • La luminosita’ di un oggetto dipende: – Dalle sue caratteristiche intrinseche – Dalla illuminazione che riceve • Variazioni su 10 ordini di grandezza Oggetti chiari, oggetti scuri • • • • • Le onde EM interagiscono con le nuvole elettroniche delle molecole degli oggetti Fotoni di determinate l vengono assorbiti e/o emessi quando provocano/derivano da transizioni elettroniche Sostanze diverse assorbono luce di diverse l Illuminando un oggetto con una luce bianca, l’oggetto riflettera’ solo la luce che non assorbe (quindi una sostanza che assorbe la luce a brevi l apparira’ rossa) Un oggetto che assorbe tutta la luce che riceve appare nero Contrasto oggetti-sfondo • Caratteristiche del segnale visivo che vengono percepite: – Contrasti di luminosita’ – Movimento coerente – Differenze di colore • L’immagine visiva NON e’ una fotografia ma un segnale molto piu’ elaborato! Il globo oculare • Tessuti trasparenti per facilitare il passaggio della luce • Fondo scuro per evitare luce riflessa (degrada la qualita’ dell’immagine) • Diaframma regolabile per dosare la luce • Lente con rifrazione variabile • Sistema di pulizia della superficie (lacrime) • Meccanismi di posizionamento (muscoli extraoculari) • Segnale in uscita: nervo ottico Formazione dell’immagine retinica • Il sistema di lenti dell’occhio forma una immagine invertita e rimpicciolita degli oggetti sulla retina • Il funzionamento e’ simile alle lenti di una macchina fotografica • Distanza focale del cristallino a riposo: 17 mm La retina • La retina e’ un foglio che ricopre la superficie posteriore dell’occhio • Punto di uscita del nervo ottico: punto cieco • Regione di massima acuita’: fovea Anatomia della retina Luce • La retina contiene tre strati di cellule: 1. Fotorecettori 2. Cellule bipolari 3. Cellule gangliari • L’epitelio pigmentato assorbe la luce che ha attraversato I fotorecettori • Le cellule orizzontali e amacrine modulano il segnale visivo trasmesso dai fotorecettori • Gli assoni delle cellule gangliari formano il nervo ottico Coni e bastoncelli • I fotorecettori della retina si dividono in coni e bastoncelli • I bastoncelli sono piu’ sensibili dei coni alla luce ma piu’ lenti e (nei mammiferi) non distinguono le l • Nei primati i coni sono di tre tipi: – Blu (short l) – Verdi (medium l) – Rossi (long l) Visione tricromatica • • • I coni blu hanno una sensibilita’ inferiore a quella dei coni verdi e rossi Il confronto tra lo stimolo che arriva ai tre tipi di coni consente di differenziare I vari colori Diverse miscele di l danno la stessa sensazione La percezione del colore • L’attivazione di un tipo di cono non da’ di per se’ la percezione del colore • Attivando in uguale misura tutti I coni si ottiene grigio o bianco • Il colore risulta dal contrasto tra I segnali dei tre coni • Il sistema RGB si basa sui tre colori primari dei coni • Il sistema Lab si basa sui tre gradienti primari della retina (luminanza-rosso/verdegiallo/blu) Anomalie della visione cromatica • Protanopia: assenza di pigmento rosso • Deuteranopia: assenza di pigmento verde • Tritanopia: assenza di pigmento blu • I numeri di Ishihara vengono usati come test per le anomalie della visione cromatica Proporzioni relative dei fotorecettori • 1 retina: •120 x 106 bastoncelli •6 x 106 coni • Schermo di questo computer: •106 pixel • Le proporzioni relative di coni e bastoncelli variano nelle regioni della retina • Nella fovea non ci sono bastoncelli • coni S: 3-5% dei coni foveali, 15% dei coni ai lati della fovea e 8% nelle altre regioni • coni M e L: difficili da distinguere - proporzioni variabili anche tra individui di una stessa specie La fovea • La fovea e’ la regione della retina con maggiore acuita’ visiva e migliore percezione dei colori • La fovea pero’ non funziona quando l’illuminazione e’ scarsa! • A livello della fovea ci sono solo fotorecettori - gli strati superiori sono spostati di lato Istologia dei fotorecettori Segmento esterno Segmento esterno Segmento interno Segmento interno Terminale sinaptico (sferula) Terminale sinaptico (pedicello) La fototrasduzione • • • • • • • • I dischi del segmento esterno contengono la proteina rodopsina (108 copie/bastoncello) Recettore a serpentina 7TM accoppiato ad una proteina G Retinale (derivato dalla vitamina A) legato alla rodopsina in forma 11-cis cis-retinale + fotone di luce visibile = trans-retinale (35 fs) cambiamento conformazionale della rodopsina. – Rodopsina – (altri intermedi metastabili) – Metarodopsina II – Opsina+retinale (alcuni ms) Circa il 50% dei fotoni che attraversano un fotorecettore vengono assorbiti L’efficienza della trasduzione e’ del 67% 1/3 dei fotoni vengono segnalati con una risposta del recettore La cascata della fototrasduzione • • Al buio, nei fotorecettori e’ fortemente attiva una guanilato ciclasi, che produce tonicamente cGMP Il cGMP mantiene aperti i canali CNG che portano una corrente cationica • • La metarodopsina II attiva la trasducina (proteina G trimera, Gt) Gta stimola una GMP ciclico fosfodiesterasi (cGMP-PDE) La luce riduce la concentrazione di cGMP e CHIUDE I canali ionici CNG La corrente al buio Buio Luce Stimolo (luce) Segmento esterno Corrente attraverso la membrana del segmento esterno Potenziale di membrana • • • • • Al buio I fotorecettori sono depolarizzati (Vz=-35/-40 mV) per la presenza di una corrente entrante stazionaria (corrente al buio) La luce chiude la corrente entrante e iperpolarizza i fotorecettori (-70 mV) La PDE e’ un’enzima molto attivo (reazione limitata dalla diffusione del cGMP) L’attivazione di 1 molecola di rodopsina idrolizza fino a 105 molecole di cGMP Quando tutti i canali sono chiusi la risposta non puo’ piu’ aumentare (saturazione) I canali cGMP-sensibili Canali cGMPsensibili • Canali cationici nonselettivi localizzati nel segmento esterno • La corrente stazionaria portata da questi canali innalza la [Na+] citoplasmatica dei fotorecettori Corrente al buio Canali selettivi per il K+ Na/K-ATPasi • Nel segmento interno una Na/K-ATPasi elimina il Na+ e aumenta il K+ citoplasmatici • Canali selettivi per il K nel segmento interno chiudono il circuito L’adattamento alla luce • Entrambi i fotorecettori adattano alla luce in pochi minuti • I coni adattano piu’ in fretta dei bastoncelli e per intensita’ luminose piu’ elevate Adattamento della fototrasduzione nel segmento esterno Il Ca2+ inibisce la guanilato ciclasi Il Ca2+ viene rimosso continuamente da uno scambiatore Na+/K+-Ca2+ Il Ca2+ inibisce la rodopsina chinasi che inattiva la metarodopsina II [Ca2+] elevata (0.5 mm) Riduzione di [Ca2+] = aumento di [cGMP] L’affinita’ dei canali CNG al cGMP e’ diminuita da Ca/ CaM I canali CNG sono permeabili al Ca2+ Uno stimolo luminoso prolungato riduce la [Ca2+] Adattamento della fototrasduzione nel segmento esterno • Adattamento varia la sensibilita’ della risposta – Bastoncello adattato al buio: 1 fotone chiude il 5% dei canali CNG in 1s – Saturazione dei bastoncelli: 10000 fotoni/s – Nei coni non si ha quasi mai saturazione completa – Dopo adattamento: canali CNG > aperti ma <sensibili – Legge di Weber • Adattamento varia anche la cinetica della risposta – Rodopsina chinasi inattiva la rodopsina in alcuni ms – Metarodopsina II stabile per vari secondi Adattamento Intensita’ percepite Aumento di luminosita’ Adattamento della sensibilita’ Differenze nella trasduzione nei coni e nei bastoncelli Bastoncelli: • maggiore superficie fotosensibile • maggiore amplificazione • Interazione rodopsina-trasducina piu’ rapida • Canali CNGA1/CNGB1a Coni: • Canali CNGA3/CNGB3 • Isoforme diverse di: – Pigmento – trasducina – PDE La risposta dei coni e’ piu’ rapida – arrestina (50-55 Hz invece di 10-12) ma meno • Maggiore [RGS9] sensibile I pigmenti fotosensibili • Sia I coni che I bastoncelli usano l’isomerizzazione del retinale • Nei coni le proteine sono dette conopsine o iodopsine e sono simili alla rodopsina ma con proprieta’ spettrali diverse • Le opsine verde e rossa sono molto simili tra loro: duplicazione genica relativamente recente (primati) Potenziale di recettore Segmento esterno Terminale sinaptico • L’assorbimento di uno o piu’ fotoni nel segmento esterno produce iperpolarizzazione • Le dimensioni dei fotorecettori (70-100 mm) consentono la propagazione passiva dei segnali di potenziale dal segmento esterno a quello interno e al terminale sinaptico • I fotorecettori non generano potenziali d’azione ma solo potenziali di recettore locali Segregazione del Ca2+ • Segmento esterno Segmento esterno: •Ca2+ implicato nell’adattamento della cascata di fototrasduzione •Ingresso: CNG •Uscita: NCKX • Terminale sinaptico Terminale sinaptico: •Ca2+ implicato nel rilascio sinaptico di neurotrasmettitore •Ingresso: canali voltaggio-dipendenti (+CNG nei coni) •Uscita: PMCA Terminale sinaptico • • • • • • Contiene vescicole di glutamato legate ad un nastro sinaptico di natura proteica Sinapsi chimica con canali per il Ca2+ attivati dalla depolarizzazione Canali di tipo L: Cav1.3 nei coni, Cav1.4 nei bastoncelli Rilascia tonicamente trasmettitore al buio (-35 /-40 mV) La luce riduce il rilascio di neurotrasmettitore (iperpolarizzazione) Nei coni una parte del rilascio e’ voltaggio-indipendente Le sinapsi dei bastoncelli Sferule dello strato plessiforme esterno Sinapsi a triade: 1 elemento presinaptico (bastoncello) contatta due elementi postsinaptici (cellula orizzontale, giallo, e cellula bipolare, rosso) Le sinapsi dei coni Pedicelli dello strato plessiforme esterno Due tipi di contatti con le cellule bipolari: Contatti invaginanti (rosso) e contatti appiattiti (blu) Sinapsi tra fotorecettori • I terminali di fotorecettori adiacenti sono collegati da sinapsi elettriche (gap junctions) • I coni blu non hanno questo tipo di sinapsi • A cosa servono questi contatti? • Ipotesi: il segnale dei coni e’ piu’ veloce, quello dei bastoncelli piu’ sensibile: accoppiamento da’ alle cellule bipolari un segnale veloce e sensibile Le cellule bipolari • Il segnale luminoso viene comunicato attraverso il terminale sinaptico dal fotorecettore alle cellule bipolari • Per I bastoncelli si ha una notevole convergenza (15-30 bastoncelli/cellula bipolare) • Il trasmettitore rilasciato e’ il glutamato • Il glutamato viene rilasciato al buio e il suo rilascio viene ridotto dalla luce Cellule ON e OFF • • • • Se si illuminano i fotorecettori direttamente in contatto con una cellula bipolare questa puo’ dare due diverse risposte: – Risposta OFF: la cellula si iperpolarizza – Risposta ON: la cellula si depolarizza!! Le cellule bipolari OFF hanno un recettore eccitatorio per il glutamato Le cellule bipolari ON hanno un recettore INIBITORIO per il glutamato Nei mammiferi tutte le cellule bipolari accoppiate ai bastoncelli sono ON Recettori per il glutamato • • • • Cellule orizzontali: recettori AMPA/kainato Cellule bipolari-OFF: recettori di tipo AMPA/kainato Cellule bipolari-ON recettori metabotropici inibitori (mGluR6) Non e’ chiaro quale sia il secondo messaggero utilizzato dalle CB-ON Le cellule bipolari • • • • • • • Postsinaptiche ai fotorecettori Presinaptiche alle cellule gangliari e amacrine Possono generare potenziali d’azione dovuti al Ca2+ Sia le sinapsi con I fotorecettori che con le cellule gangliari sono sinapsi a nastro glutamatergiche Le cellule bipolari (CB) formano sinapsi glutamatergiche con le cellule gangliari (CG) La sinapsi con le cellule gangliari e’ fasica (depressione post-stimolo) Le cellule gangliari hanno solo recettori eccitatori La via diretta • Il circuito piu’ semplice nella retina contiene: – uno o piu’ fotorecettori – Una o piu’ CB – Una CG • In questo circuito lo stimolo efficace e’ una luce (o un’ombra) che colpisce una regione ristretta della retina • L’entita’ della convergenza e’ estremamente variabile Tipi di cellule bipolari • Nei primati ci sono 10 tipi di CB • Le cellule nane (midget cells) della fovea non hanno convergenza: ricevono input da 1 solo cono e trasmettono output a 1 cellula gangliare • Massima discriminazione spaziale Campi recettoriali • Un singolo cono risponde alla luce su una piccola frazione di retina Contrasto centro-periferia • Una CB che raccoglie le risposte di piu’ fotorecettori dovrebbe avere meno sensibilita’ spaziale • Quindi illuminando una regione via via piu’ estesa della retina si dovrebbe avere una risposta via via maggiore della CB • In realta’ le CB hanno una risposta bifasica (centro-periferia) La sinapsi dei fotorecettori • I fotorecettori rilasciano glutamato a riposo • La luce iperpolarizza I recettori… • Depolarizza le CB-ON dei bastoncelli e dei coni… • Iperpolarizza le CB-OFF dei coni… • Iperpolarizza le cellule orizzontali Le cellule orizzontali Illuminando un fotorecettore vengono iperpolarizzate le cellule orizzontali che esso contatta • • • • • Cellule inibitorie (GABAergiche o glicinergiche) Assone corto o inesistente Arborizzazione dendritica estesa nel piano dello strato plessiforme esterno Ricevono input eccitatorio dai fotorecettori Accoppiate tra loro con gap junctions (chiuse dai recettori D1 dopaminergici) La via indiretta • I fotorecettori che contattano direttamente una CB (in blu) producono la risposta principale (centro) ON o OFF • I fotorecettori (in bianco) che contattano le cellule orizzontali connesse alla via diretta (in ocra) inibiscono I fotorecettori della via diretta e quindi hanno un effetto contrario sulla CB (eccitatori su CB-OFF e inibitori su CBON) • Si genera cosi’ opposizione centro-periferia Campi recettoriali delle cellule gangliari La visione a colori • I meccanismi di opposizione centro-periferia funzionano per la luminanza del segnale acromatico ma anche per I colori • Due tipi di opposizione cromatica: – Rosso-verde – Blu-giallo Da dove viene il giallo? • Il giallo deriva dalla fusione dei segnali dai coni verdi e rossi • I coni blu hanno vie distinte fino al SNC • I coni rossi e verdi no (evoluzione piu’ recente) La sinapsi dello strato plessiforme interno • • • • • • • CG solo postsinaptiche CB, cellule amacrine hanno sinapsi bidirezionali La via diretta CB-CG viene pesantemente modulata Per I bastoncelli non c’e’ una via diretta nei primati Cellule bipolari: 10 tipi diversi Cellule gangliari: 20 tipi diversi Cellule amacrine: 30 tipi diversi • • Via diretta: sinapsi CB-CG Via indiretta: sinapsi CB-cellule amacrine La sinapsi dello strato plessiforme interno • • • • Architettura molto complessa Ruolo delle cellule amacrine spesso ancora poco chiaro e variabile a seconda della specie Nel coniglio, una classe di cellule amacrine fornisce segnali su oggetti in movimento Nel gatto, l’elaborazione del movimento avviene nel SNC Neurotrasmettitori nella retina • La via diretta fotorecettoreCB-CG e’ glutamatergica • L’output primario delle cellule orizzontali e’ GABAergico • Altri fotorecettori e modulatori sono stati osservati, per molti dei quali non e’ ancora nota la funzione Differenze tra specie • La retina dei vertebrati non mammiferi e’ solitamente adattata per la visione diurna (problema di saturazione) • La retina dei roditori e dei carnivori e’ adattata per la visione notturna (predominanza dei bastoncelli, problema di sensitivita’) • La retina della tupaia e’ composta quasi esclusivamente da coni • I primati sono adattati per la visione del colore e I dettagli fini Le cellule gangliari • • • • • • • • 1 milione di cellule gangliari/125 milioni di fotorecettori Generano spikes Unico output della retina Le CG-P (b) ricevono segnali lenti e tonici da campi recettoriali molto piccoli (80%) Le CG-M (a) ricevono segnali rapidi e fasici da ampie regioni retiniche (<10%) Altre cellule sono classificate come nona, non-b I campi recettoriali di entrambe le classi ricoprono tutta la retina e sono piu’ piccoli nella fovea Tutte le classi di CG possono essere ON o OFF Cellule gangliari beta • Trasformano in scarica di potenziali d’azione il potenziale graduale delle CB • Moderata convergenza • Opponenza di luminanza o cromatica • Segnale: contrasto tra punti adiacenti (selettivita’ ai bordi) Cellule gangliari alfa • La CG alfa ricostruita riceve input da 150 CB, 250 cellule amacrine IIB e > 1000 cellule amacrine di altro tipo • Ciascuna CB riceve input diretti da 10-15 coni • Le CG alfa riflettono l’illuminazione generale di ampie regioni della retina CG sensibili al movimento • Alcune CG rispondono soltanto a stimoli puntiformi che si muovono in una data direzione!! • Non e’ chiaro se esistano nell’uomo • Responsabili di questo comportamento sono le cellule amacrine starburst Le cellule amacrine • 30 tipi diversi • CA glicinergiche: solitamente campo ristretto • CA GABAergiche: solitamente campo esteso • CA starburst rilasciano sia GABA che ACh: danno selettivita’ direzionale alle CG Le cellule amacrine CA-II • • • • Non esiste una via diretta ON dai bastoncelli alle cellule gangliari CA-II (glicinergiche) accoppiano le cellule bipolari CB-ON dei bastoncelli con la via diretta ON dai coni (gap junctions) e con la via OFF dei coni (glicina) nei primati la visione data dai soli bastoncelli si ha raramente – in assenza di coni si e’ legalmente ciechi, mentre in assenza di bastoncelli si hanno solo problemi di visione notturna L’accoppiamento elettrico dato dalle cellule CA-II viene modulato da cellule amacrine dopaminergiche
Scaricare