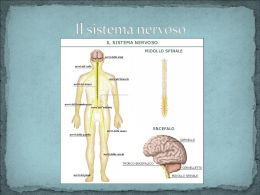
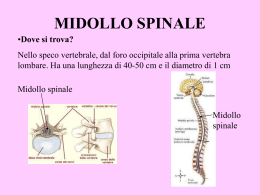
SISTEMA NERVOSO: Il SNC: “Il SNC rappresenta l'insieme delle formazioni nervose contenute all'interno della cavità cranica e del canale vertebrale” Il SNC riceve informazioni sia dall'ambiente esterno sia da quello interno, tramite fibre che portano informazioni dalla periferia (fibre afferenti del SNP) questo se ci sono speciali recettori come termocettori, chemiocettori, pressocettori, recettori ormonali, ecc. Tali informazioni raggiungono il SNC dove vengono elaboratein modo da generare risposte adeguate alla natura dello stimolo percepito, che vengono inviate alla periferia tramite fibre che escono dal SNC (fibre efferenti del SNP). Nonostante la sua complessità, il SNC risulta costituito dalla ripetizione di circuiti (catene neuronali) costituiti da: 1. NEURONE SENSITIVO, cellula ch prende contatto con la periferia dove raccoglie le informazioni 2. NEURONE EFFETTORE, cellula che invia messaggi alla periferia per realizzare contr. muscolari o attività secretorie 3. INTERNEURONE, cellula interposta tra le prime due che modula l'informazione in arrivo con il segnale in uscita, anche se non è sempre presente e ne è dimostrazione il “riflesso” Il SNC si compone di midollo spinale e encefalo Il midollo spinale: Il midollo spinale è in diretta continuazione con l'encefalo, è accolto nel canale vertebrale e ha la forma di un lungo cilindro leggermente schiacciato in senso antero-posteriore. È lungo circa 44 cm e un diametro medio di 1 cm. È collegato alla periferia tramite una doppia serie di 33 nervi spinali a disposizione metamerica, che fuoriescono dal canale vertebrale attraverso i forami intervertebrali. Ogni nervo si biforca in due radici, una anteriore e una posteriore, che sono in rapporto con il MS. 2 4 3 SEGMENTO MIDOLLARE CERVICALE NERVI CERVICALI SEGMENTO MIDOLLARE DORSALE NERVI DORSALI 1 SEGMENTO MIDOLLARE LOMBARE SEGMENTO MIDOLLARE SACRALE 1. Solco mediano anteriore 2. Solco mediano posteriore 3. Solco laterale anteriore 4. Solco laterale posteriore NERVI LOMBARI CAUDA EQUINA NERVI SACRALI NERVI COCCIGEI Figura 1 Per effetto della ascensione midollare, al termine del suo sviluppo, il MS non riempie completamente il canale midollare, ma si arresta all'altezza della seconda vertebra lombare. Questo fa si che le paia di nervi spinali che escono dai forami intervertebrali dei primi neuromeri, compiano un tragitto pressochè orizzontale mentre via via che si scende lungo il MS, i nervi raggiungono i siti di emergenza in direzione sempre più obliqua fino a che gli ultimi, hanno una decorrenza verticale formando un fascio chiamato cauda equina. Figura 2 Il MS è circondato da tre lamine connettivali concentriche, le meningi spinali. A stretto contatto con il midollo, c'è la pia madre, più esternamente l'aracnoide e infine la dura madre. Fra pia madre e aracnoide è presente uno spazio detto subaracnoideo che contiene il liqor cefalorachidiano, fra aracnoide e dura madre vi è uno spazio virtuale detto subdurale e infine tra dura madre e il periostio interno che delimita il canale midollare, vi è un altro spazio detto epidurale che contiene plessi venosi e tessuto adiposo. In superficie il MS presenta dei solchi (figura 1): solco mediano anteriore, occupato da un setto della pia madre, un solco mediano posteriore, meno accentuato, e dei solchi laterali che corrispondono all'origine apparente dei nervi spinali. Infatti solo le radici anteriori sono costituiti da assoni di neuroni effettori (che portano gli impulsi alla periferia, che innervano i muscoli scheletrici, motoneuroni somatici, oppure la muscolatura liscia e ghiandole, neuroni visceroeffettori) e che quindi originano realmente dal midollo. Quelle posteriori sono formati infatti da assoni di neuroni sensitivi posti fuori del MS, che quindi inviano al midollo le informazioni raccolte in periferia, per cui la loro origine è solo apparente. Il canale vertebrale è occupato solo centralmente dal MS che si assottiglia nel cono midollare dove le meningi spinali, accompagnandosi alla cauda equina, si fissano alla faccia posteriore del coccige tramite il legamento coccigeo. Tale legamento, la dura madre e la continuità anatomica del MS, fanno si che questo rimanga in posizione centrale nello speco vertebrale. Il liqor, inoltre, garantisce un cuscinetto protettivo da traumi e compressioni a carico del MS. Il calibro del MS non è uniforme (figura 1) ma presenta dei rigonfiamenti a livello cervicale e a livello lombare conseguenza della massiccia presenza di innervatura motrice e sensitiva degli arti superiori e inferiori. Tipi di sensibilità: Si distinguono vari tipi di sensibilità in base alla provenienza o alla direzione dello stimolo: Sensibilità esterocettiva (a sua volta divisibile in specifica e generale). Consente di percepire stimoli provenienti dal mondo esterno o metterci in relazione con esso. Se è richiesta la presenza di organi specifici come l’occhio o l’apparato uditivo, posti nella testa, si parla di sensibilità esterocettiva specifica, mentre se le informazioni provengono da recettori diffusi nella cute o nelle mucose vicine ad essa, si parla di sensibilità esterocettiva generale. Questo tipo di sensibilità si può suddividere ulteriormente in epicritica e protopatica: la prima da informazioni dettagliate sulla natira dello stimolo e sulla sede di applicazione ed è sempre di tipo cosciente. La seconda invece, fornisce informazioni più grossolane ed è spesso di tipo incosciente. Sensibilità viscerale o introcettiva origina dai visceri, è sempre di tipo protopatico, quindi non dà mai informazioni coscenti precise sullo stimolo. La componente cosciente è dolorifica mentre quella incosciente è utilizzata per i riflessi. Sensibilità propriocettiva origina da particolari recettori localizzati a livello dei muscoli scheletrici, dei tendini, dei legamenti e delle capsule articolari e permette di percepire lo stato di contrazione dei muscoli e la posizione nello spazio delle parti del corpo. Può essere protopatica o anche epicritica. Una forma particolare è quella cinestatica, che origina da un apposito apparato posto nell’orecchio interno detto apparato vestibolare o dell’equilibrio in seguito a spostamenti del capo o dell’intero corpo. Figura 3 La struttura del midollo spinale: La sostanza grigia è disposta al centro circondata dalla sostanza bianca in posizione periferica. Centralmente è percorso per quasi tutta la sua lunghezza dal canale ependimale, residuo del primitivo tubo neurale che prosegue encefalicamente aprendosi nella cavità del quarto ventricolo e contiene liquor. La sostanza grigia in sezione ha la forma di una farfalla con una parte trasversale detta commessura grigia percorsa dal canale ependimale. Le ali formano un paio di corna anteriori e uno di corna posteriori. La sostanza grigia risulta costituita da colonne di neuroni, mentre la sostanza bianca è organizzata in fasci di fibre nervose prevalentemente mieliniche. La sostanza bianca, che è a contatto con le meningi, viene suddivisa dalla sostanza grigia in porzioni dette cordoni o funicoli: Per ogni metà detta antimero, si Figura 4 distinguono un cordone posteriore (figura 3 regione rossa) tra il solco mediano posteriore e il corno posteriore, un cordone laterale (figura 3 regione gialla) tra il corno posteriore e quello anteriore e un cordone anteriore (figura 3 regione blu) tra il corno anteriore e il solco mediano anteriore. I cordoni sono formati da fasci di fibre nervose mieliniche che danno la colorazione bianca. I fasci discendenti trasportano dall'encefalo alla periferia stimoli di natura motoria da trasmettere attraverso i nervi spinali, i fasci ascendenti invece portano stimoli sensitivi dalla periferia all'encefalo. Le fibre nervose mieliniche che decorrono verticalmente e vanno a formare questi fasci(ascendenti e discendenti) sono dette fibre funicolari e possono essere distinte in endogene(nate dal midollo spinale stesso)ed esogene(nate dall'encefalo e discendenti nel midollo spinale). Vi sono poi altre fibre nervose mieliniche che a differenza delle funicolari hanno un decorso grosso modo orizzontale, queste sono le fibre commesurali e le fibre radicolari. La sostanza bianca dell'encefalo ha una posizione leggermente diversa da quella nel midollo spinale, ma la funzione è simile. Nel cervelletto e nel cervello si trova sotto la corteccia composta da sostanza grigia, ma più esternamente rispetto ai nuclei (gruppi di neuroni che costituiscono una parte della sostanza grigia dell'encefalo). Le varie porzioni in cui è distinto l'encefalo rendono individuabile vari tipi di fasci che costituiscono la sostanza bianca. Abbiamo infatti fasci ascendenti, ovvero la continuazione dei fasci ascendenti del midollo spinale, che sono destinati a terminare in corrispondenza della corteccia cerebrale; fasci discendenti, ovvero l'origine di una parte dei fasci discendenti del midollo spinale destinati a portarsi direttamente in periferia o a collegarsi con la sostanza grigia del midollo spinale. Inoltre sono da considerare i numerosissimi fasci di associazione che collegano le varie parti dell'encefalo. Nella sostanza grigia sono presenti neuroni del I tipo di Golgi, cioè caratterizzati da un lungo assone mielinico che fuoriesce dalla sostanza grigia stessa e del II tipo di Golgi, ad assone breve amielinico che rimane all’interno della sostanza grigia. Quelli del I tipo sono neuroni che si trovano nel corno anteriore e possono essere somatomotori ( motoneuroni alfa e gamma), che innervano la muscolatura striata scheletrica, e visceroeffettori (funicolari o motoneuroni beta), che innervano la muscolatura liscia dei visceri e delle ghiandole. Si tratta di neuroni multipolari che fuoriescono dal solco laterale anteriore ed entrano a far parte dei nervi spinali che quindi originano realmente dal MS. I motoneuroni si distinguono in alfa e gamma: Gli alfa hanno assoni che si dirigono verso le fibre muscolari in prossimità delle quali si ramificano ripetutamente e ogni ramificazione contrae sinapsi con una fibra muscolare (placca motrice). L’unità motoria è un numero di fibre che dipende dalla ramificazione di un singolo neurone, più piccola è l’unità motoria e più raffinato sarà il controllo muscolare e quindi il movimento. I gamma sono meno numerosi e i loro assoni innervano i sensori dello stato di contrazione e distensione muscolare informando l’asse nervoso permettendo al SNC di controllare l’attività contrattile (circuito gamma). I visceroeffettori appartengono al SNA,sono più piccoli e sono localizzati alla base del corno anteriore e nel corno laterale dove formano la cosiddetta colonna intermedio-laterale. Le cellule radicolari visceroeffettrici sono distinte in due gruppi: neuroni ortosimpatici e parasimpatici. Gli ortosimpatici sono localizzati nella colonna intermedio-laterale (dall’VIII neuromero cervicale al II neuromero lombare), mentre le cellule parasimpatiche sono ubicate nella parte sacrale del MS (dal II al IV neuromero sacrale). Gli assoni di questi neuroni fuoriescono insieme a quelli dei motoneuroni dal MS per costituire le radici anteriori dei nervi spinali ma abbandonano il nervo spinale per raggiungere i gangli simpatici provvisti di neuroni effettori viscerali. Sono quindi detti fibre pregangliari.Le cellule localizzate nelle corna posteriori ricevono informazioni dai sensori periferici per la sensibilità somatica e sono, quindi detti neuroni somatosensitivi, o per la sensibilità viscerale detti neuroni viscerosensitivi. Le cellule funicolari sono dette così in quanto emettono assoni che entrano nella sostanza biancadividendosi in un ramo ascendente e uno discendente per rientrarepoi nella sostanza grigia e mettendo quindi in comunicazione diverse parti dell’asse nervoso. RIASSUMENDO Nella sostanza grigia le cellule sono raggruppate in colonne o nuclei per una certa lunghezza del ms TESTA DELLE CORNA ANTERIORI neuroni somatomotori - innervazione della muscolatura scheletrica BASE DELLE CORNA ANTERIORI E CORNA LATERALI neuroni visceroeffettori - innervazione muscolatura liscia e vari organi CORNA POSTERIORI neuroni somatosensitivi e viscerosensitivi Vie motorie piramidali e extrapiramidali: La via piramidale è mononeuronica e molto rapida; la via extrapiramidale è formata da catene di neuroni ed è più lenta. Tutte e due convergono nei motoneuroni che si trovano nelle corna anteriori del midollo spinale. La stessa cosa avviene per i nuclei motori dei nervi cranici situati nel tronco encefalico, che ricevono vie corticonucleari mononeuroniche (omologhe alle vie piramidali) e vie polineuroniche (omologhe a quelle extrapiramidali). Per unità motoria si intende la cellula motrice, cellula radicolare anteriore, e le fibre muscolari da essa innervate. Via piramidale: ha origine dalla corteccia motoria, passa per una porzione del bulbo chiamata piramide, da cui prende il nome, ed arriva al midollo spinale. Serve per l'esecuzione del movimento. La via piramidale nell'Uomo è suddivisa in: • fascio piramidale crociato • fascio piramidale diretto. Esistono fibre piramidali molto lunghe, come quelle che vanno dalla corteccia frontale ai mielomeri sacrali-coccigei. Più corte sono, ovviamente, quelle che si interrompono ai mielomeri cervicali, toracici, lombari. Le fibre aventi come bersaglio i motoneuroni sono le cosiddette fibre discendenti, e provengono dalla corteccia cerebrale.Una delle principali vie discendenti è la via piramidale (detta anche via cortico-spinale), così chiamata perché passa attraverso le piramidi, formazioni situate a livello del bulbo. Tale via presenta due porzioni: - Via cortico-nucleare (che si ferma nel bulbo), destinata ai nuclei dei nervi cranici - Via cortico-spinale propriamente detta, che giunge fino agli ultimi neuromeri del midollo spinale. Ne consegue che le fibre di quest'ultima via sono molto lunghe. A livello delle piramidi del bulbo, gran parte delle fibre della via piramidale si incrocia e decorre nei cordoni laterali del midollo spinale, ma vi è anche una piccola porzione di fibre (dal 5% al 15%) che rimane omolaterale. Queste sono state scoperte relativamente di recente, osservando che, in caso di lesioni della zona corticale (per esempio nella parte destra) che provocano una paralisi, si ha un parziale recupero del movimento. Infatti, le fibre non crociate mantengono la loro funzionalità e sostituiscono in parte quelle danneggiate. Questo fenomeno ha un andamento soggettivo, in quanto il numero di fibre che non si incrociano varia da individuo a individuo. Il contingente di fibre destinato al cordone anteriore non si incrocia nel bulbo, ma si incrocia appena arriva a destinazione; dunque, questo fascio è funzionalmente crociato e si estingue a livello dei primi mielomeri toracici, in quanto è deputato all'innervazione dei muscoli rotatori, estensori e flessori del capo. Come sappiamo, i vasi del cervello, come quelli del cuore, sono vasi terminali, per cui se si verifica una lesione a carico di uno di questi vasi, la parte di tessuto che si trova a valle della stessa va incontro a infarto o necrosi ischemica: mancando il sangue, quella zona di encefalo muore e viene sostituita con tessuto gliale. In questo caso le fibre si interrompono a livello della capsula interna e si avrà la lesione delle vie motorie. Se la lesione è elettiva per le vie piramidali, in una zona precisa della capsula interna, per esempio nella parte destra, i motoneuroni della parte sinistra del midollo spinale non riceveranno più gli impulsi; vengono colpiti i motoneuroni della parte sinistra perché le fibre si incrociano a livello del bulbo: la risposta è dunque eterolaterale. Via extrapiramidale: ha origine dalla corteccia ed arriva al midollo spinale, dopo essersi interrotta in vari nuclei sottocorticali. È finalizzata alla regolazione del movimento. Esistono diversi tipi di unità motrici in relazione alle fibre muscolari che le formano e al tipo di motoneurone che le innerva. A seconda del tipo di muscolo ci possono essere unità motorie formate da centinaia o migliaia di fibre e altre formate da poche fibre, come ad esempio i muscoli della mano, della lingua, che servono per modulare le parole e quelli degli occhi. Questi ultimi sono importanti per una perfetta sovrapposizione dei globi oculari, in mancanza della quale abbiamo disturbi funzionali ed antiestetici come lo strabismo e la diplopia (percezione simultanea di due immagini di uno stesso oggetto) Le vie extrapiramidali hanno diversi fasci, tra cui il più importante è quello "reticolo-spinale", che ha due contingenti: uno decorre nel cordone laterale e si chiama "fascio bulbo- reticolare" perché proviene dalla sostanza reticolare del bulbo; l'altro decorre nel cordone anteriore e proviene dalla sostanza reticolare del mesencefalo e del ponte. Tali fasci sono estremamente importanti. Le vie reticolo-spinali anteriori, infatti, sono quelle che eccitano gli estensori regolando il tono estensorio; di conseguenza, in caso di danno del midollo spinale, si può avere una lesione di tali vie che provoca lo sviluppo di una paralisi spastica in flessione, impedendo così la deambulazione. Un altro fascio molto importante delle vie extrapiramidali è quello "vestibolo-spinale". Anche questi fasci sono duplicati nel cordone anteriore ed in quello posteriore, sono dunque diversi contingenti di fibre, ma con la stessa funzione. La grande importanza dei fasci vestibolo-spinali può essere constatata, per esempio, osservando alcuni movimenti del gatto e dell'uomo. Infatti, quando il gatto è intenzionato a mangiare un canarino, guarda la gabbia del medesimo alzando la testa, ponendo le zampe posteriori in posizione flessa e quelle anteriori in posizione estesa. Nel momento in cui il gatto intende saltare addosso alla sua preda, abbassa la testa e conseguentemente si ha una flessione delle zampe anteriori, nonché un'estensione di quelle posteriori. La stessa cosa accade all'uomo nel momento in cui, sporgendosi, abbassa la testa: egli non cade, perché la testa governa i muscoli estensori. Da questi due esempi, possiamo evincere che i fasci vestibolo-spinali, importantissimi, rappresentano il terzo sistema di regolazione del movimento (insieme ai fasci tetto-spinali ed al sistema della sensibilità propriocettiva cosciente), essendo essi coinvolti nei riflessi di raddrizzamento sulla base dei movimenti del capo. I riflessi spinali Il midollo spinale non è una semplice stazione di transito dei segnali in ingresso ed in uscita ma presenta strutture capacità di generare semplici risposte proprie (riflessi) ed anche eccitazioni cicliche e sincronizzate che provocano movimenti ripetitivi anche di una certa difficoltà. Cercheremo quindi di approfondire le strutture che sono alla base delle funzioni ed i loro meccanismi di azione partendo ancora dal muscolo che contiene i recettori specifici in grado di inviare segnali al midollo per passare poi ad analizzare le funzioni del midollo spinale propriamente detto. I riflessi spinali, che costituiscono il primo livello dell'organizzazione motoria, si basano sulla risposta diretta a stimoli recettoriali. I recettori specializzati del muscolo sono i fusi neuromuscolari (vedi struttura del fuso neuromuscolare) e gli organi muscolo tendinei del Golgi (vedi struttura dell'organo del Golgi). Entrambi questi recettori sono connessi con fibre nervose rapide, rispettivamente di tipo Ia e Ib, che inviano segnali sia a livello spinale che a livelli superiori. La conformazione e la posizione di questi due tipi di recettori permette di controllare la tensione e la lunghezza del muscolo in modo molto preciso (vedi recettori). I fusi in particolare presentano non sono dei semplici "trasduttori del segnale" ma svolgono una funzione più articolata di amplificazione o smorzamento in quanto la loro risposta può essere controllata da livelli superiori attraverso l'attività dei motoneuroni gamma. Questo tipo particolare di neurone motorio ha connessioni solo con le fibre intrafusali (non contribuisce quindi direttamente al movimento) e presenta caratteristiche funzionali diverse, si possono infatti riconoscere m. gamma statici e dinamici che agiscono in modo nella modulazione delle risposte del fuso in condizioni stabili e nelle transizioni. A seguito dell'eccitazione dei due tipi di neuroni è possibile migliorare la performance del sistema fusale nel monitorare la lunghezza del muscolo in sé oppure controllare piccole e rapide fluttuazioni. La presenza dei m.gamma permette anche al sistema nervose di controllare la sensibilità dei fusi anche nel corso della contrazione attiva. Infatti la contemporanea attivazione di m. alfa e gamma consente al fuso di continuare ad informare il sistema nervose centrale anche durante l'accorciamento del muscolo (attivazione alfa-gamma) Vari studi condotti da Valbo e coll. Hanno dimostrato che senza la presenza dell'attivazione gamma nel corso del movimento la scarica afferente cesserebbe e la possibilità di controllo del movimento sarebbe gravemente ridotta. E' stato inoltre dimostrato che l'attivazione gamma non è solo dipendente dalla quantità di attivazione alfa, ovvero dalla forza di contrazione sviluppata, ma anche dalla complessità del compito svolto. Infatti i m. gamma sia statici che dinamici vengono attivati in misura via via maggiore mano a mano che aumenta la velocità e la difficoltà del movimento. (vedi attività fusimotoria). Dopo aver considerato le caratteristiche funzionali del fuso neuromuscolare vediamo come il sistema nervoso lo utilizza come struttura base della forma più semplice di movimento, il riflesso. Il riflesso più importante e certamente il più studiato è il riflesso da stiramento o miotatico descritto verso la fine dell'800 da Sherrington il quale, osservando come il movimento venisse abolito dalla sezione delle radici dorsali o ventrali, dimostrò l'origine spinale del riflesso. Il r. da stiramento viene evocato dall'allungamento del muscolo e provoca una risposta contrattile che tende e a ridurre lo lunghezza del muscolo stesso. La risposta presenta due componenti, una fasica di breve durata evocata dal rapido cambiamento di lunghezza ed una tonica indotta anche da lente variazioni del muscolo. L'attività riflessa non riguarda solo il muscolo stirato che si contrae ma anche il muscolo antagonista che si rilascia (fenomeno detto innervazione reciproca vedi oltre). Questa risposta avviene con una latenza molto bassa che è spiegabile solo attraverso la presenza di una sinapsi diretta tra fibra afferente e neurone motore (riflesso monosinaptico) ed è proporzionale all'intensità dello stiramento ed alla velocità con cui viene provocata. E' stato recentemente postulato che ogni fibra Ia afferente sia in grado di attivare tutti i motoneuroni che innervano il muscolo allungato ed i mm. sinergici. Lo schema di funzionamento del riflesso è qui mostrato nel dettaglio. (vedi riflesso da stiramento). L'intensità della risposta può anche essere modulata dal controllo sovraspinale attraverso la contrazione delle fibre intrafusali stimolate dai gamma motoneuroni che permettono di migliorare la risposta alle variazioni rapide di lunghezza con una contrazione altrettanto efficace. L'attività statica induce invece una risposta continua che è nota con il termine di tono muscolare che definisce la forza con la quale il muscolo reagisce all'allungamento, ovvero la sua rigidezza. Alla rigidezza dovuta ai componenti elastici si somma una componente nervosa causata dal riflesso da stiramento. Il tono contribuisce al mantenimento della postura riducendo le oscillazioni antero-posteriori sia nella stazione eretta che durante la locomozione. Inoltre queste proprietà che conferiscono al muscolo qualità elastiche lo rendono simile ad una molla e contribuiscono a rendere il movimento scorrevole.Un eccesso di tono muscolare, o un eccessivo guadagno del sistema di controllo fusimotorio, deriva da lesioni delle vie discendenti che fanno mancare una adeguata inibizione dei m.gamma provocando una ipertonia muscolare la cui forme più comuni sono la spasticità (resistenza eccessiva allo stiramento) ed il clono (rapida successione di flessioni e estensioni parziali). Il riflesso flessorio consiste nella flessione simultanea delle tre maggiori articolazioni di un arto ed è evocato di solito da stimoli di natura dolorifica ma può derivare anche da latri stimoli (tattile, cutanei) . Di norma nel soggetto integro soli i recettori dolorifici attivano il riflesso ma in condizioni di spinalizzazione (interruzione del controllo superiore sul midollo) la risposta flessori si evidenza anche con modesti stimoli tattili. Quindi esiste una azione inibitoria sulla via spinale del riflesso che parte dalla sostanza reticolare del tronco encefalico. Nella paraplegia da interruzione del midollo oltre alla flessione si manifestano anche altre risposte di tipo vegetativo (sudorazione, piloerezione, arrossamento della cute). Il riflesso flessorio è quindi un riflesso polisinaptico mediata da diverse afferenze dei gruppi II III e IV (definite in modo globale Flexor Reflex Afferents) che attivano vari interneuroni spinali introducendo quindi un ritardo di trasmissione nella risposta (latenza centrale di 6-7 msec più del doppio del riflesso monosinaptico) ed un certo grado di aspecificità (la flessione riguarda vari gruppi muscolari) con un'attivazione che si mantiene ben oltre la durata dello stimolo. Il campo motore del riflesso flessorio è molto esteso ma l'entità relativa dell'attivazione dei diversi gruppi muscolari dipende dalla sede di stimolazione. Ad esempio se lo stimolo è applicato alla pianta del piede si contraggono vivacemente i flessori del piede e della gamba e più moderatamente quelli della coscia. Spesso la flessione omolaterale rispetto alla sede dello stimolo si accompagna ad una estensione controlaterale (riflesso estensorio crociato) che si accompagna anche all'inibizione reciproca dei motoneuroni antagonisti. Il riflesso plantare si produce strisciando con un oggetto smusso la parte laterale della pianta del piede (vedi riflesso plantare), ciò provoca una risposta flessoria delle dita. Se esiste invece una lesione delle vie cortico-spinali la reazione si capovolge e si osserva una flessione dorsale delle dita e la loro divaricazione (riflesso di Babinski). Questa evidenza indica che gli interneuroni che mediano la risposta riflessa sono sotto il controllo corticale (in condizioni normali sono facilitati dalle vie corticospinali) la cui assenza modifica il comportamento del circuito neurale spinale invertendo il movimento prodotto dalla stimolazione recettoriale. La corteccia cerebellare Il cervelletto, situato nella regione postero-inferiore della cavità cranica, tra il cervello e il midollo allungato, è diviso in due emisferi cerebellari, uniti al verme cerebellare. I peduncoli cerebellari collegano il cervelletto alle tre parti del tronco cerebrale (mesencefalo, ponte di Varolio e bulbo o midollo allungato), mentre il tentorio lo collega al cervello. La corteccia cerebellare è costituita da tre strati di sostanza grigia: uno strato esterno o molecolare, uno intermedio e uno interno o granulare. Nella corteccia cerebellare si ramifica la sostanza bianca (arbor vitae, albero della vita) che contiene nuclei di sostanza grigia. Il cervelletto (che costituisce il 10% del peso encefalico) è l'organo deputato a coordinare i movimenti muscolari, presiede al senso dell'orientamento del corpo nello spazio e al mantenimento dell'equilibrio. Le lesioni che possono colpirlo sono di varia natura (congenita, infiammatoria, traumatica, neoplastica) e si manifestano con tremori, disturbi della parola, della scrittura, dell'equilibrio e altri. La corteccia cerebellare è stratificata alla superficie del cervelletto, coprendo in maniera uniforme la sostanza bianca, che così risulta centrale. E'identica in tutta la superficie dell'organo ed è composta da tre strati di neuroni che, dall'esterno all'interno, sono: lo strato molecolare, lo strato gangliare o delle cellule del Pùrkinje, lo strato granulare. Poichè le cellule di Pùrkinje sono i neuroni più caratteristici del cervelletto, iniziamo la descrizione da questo strato. Le cellule di Pùrkinje, o cellule gangliari, sono neuroni con un voluminoso soma a forma di pera disposti tutti in un unico strato fra lo strato molecolare e quello granulare. Dalla base del soma nasce il neurite che penetra nello strato granulare, lo attraversa tutto e si dirige ai nuclei centrali del cervelletto, dove di solito termina. Dal collo del soma piriforme prendono origine due o tre grossi dendriti che penetrano nello strato molecolare, diretti verso la superficie. I dendriti si ramificano alcune volte costituendo una spalliera e tutti i rami assumono una direzione comune verso la superficie della corteccia, cioè risultano paralleli tra loro. Ma non basta: sono anche tutti disposti in un unico piano, per cui una cellula di Pùrkinje con i suoi dendriti assomiglia ad un candelabro ebraico. Anche questo non basta: i piani delle spalliere dendritiche di tutte le cellule di Pùrkinje sono paralleli tra loro, perpendicolari alla superficie della corteccia e perpendicolari al piano che forma l'asse di ogni lamella cerebellare. I rami dendritici delle spalliere sono ricchi di piccole estroflessioni (le spine) su cui poggiano le sinapsi di altri neuroni. Infatti con le cellule di Purkinje prendono rapporto sinaptico i neuriti delle cellule dei canestri dello strato molecolare (a livello del soma), i neuriti dei granuli dello strato molecolare (si appoggiano sulle spine) e i neuriti di neuroni del nucleo olivare bulbare, che si arrampicano sulle spalliere (fibre rampicanti) e che pure si appoggiano sulle spine. Lo strato molecolare è formato da interneuroni di associazione di varia forma e dimensioni, piuttosto scarsi (da cui il nome molecolare dello strato), tra i quali si riconoscono, in prossimità dello strato di Pùrkinje, le cellule dei canestri. Questi neuroni emettono un neurite che è sempre diretto perpendicolarmente al piano della lamella cerebellare (cioè parallelamente ai piani delle spalliere dendritiche), e ogni volta che incontra il soma di una cellula di Pùrkinje, si ramifica attorno ad esso formando il canestro e le sinapsi. Ogni cellula dei canestri contatta una decina di cellule di Pùrkinje. Nello strato molecolare sono presenti anche i neuriti dei granuli, che qui salgono e si dividono a T, formando le fibre parallele, così dette perchè sono tutte parallele tra loro, parallele alla superficie della corteccia e perpendicolari ai piani delle spalliere: le fibre parallele prendono contatto sinaptico con le spine dei dendriti delle cellule di Purkinje. Ancora, nello strato molecolare si trovano parte dei dendriti delle cellule di Golgi dello strato granulare, con i quali prendono contatto saempre le fibre parallele. Per finire lo strato granulare è formato dai granuli, piccoli neuroni dal soma sferico, con quattro o cinque dendriti terminanti con piccole ramificazioni "ad artiglio" e il neurite che sale nello strato molecolare per formare una fibra parallela. Tra i granuli, che ricevono dall'esterno le fibre muscoidi con tutte le informazioni da integrare, sono sparsi i grandi granuli o cellule di Golgi, il cui neurite si ramifica ripetutamente nello stesso strato granulare e contatta i piccoli granuli, mentre i dendriti si ramificano in tutte le direzioni, salendo anche nello strato molecolare. Gli unici neuriti che abbandonano la corteccia cerebellare sono quelli delle cellule di Pùrkinje, pochi dei quali però abbandonano il cervelletto, in quanto in gran parte si portano ai nuclei centrali. Sistema nervoso simpatico (neurovegetativo) Rappresenta quella parte del sistema nervoso deputata alla regolazione delle funzioni viscerali che normalmente sfuggono al controllo della volontà, come il controllo della muscolatura liscia dei vasi e dei diversi visceri, la secrezione ghiandolare e il controllo della frequenza cardiaca. Viene diviso in due grandi componenti, l’ortosimpatico, o simpatico propriamente detto, e il parasimpatico. Dal punto di vista anatomico, le differenze tra i due componenti risiedono nella localizzazione dei centri effettori viscerali, che per il primo sono ubicati a livello toraco-lombare del MS da T1 a L3, e per il secondo a livello dei nervi encefalici siti nel tronco encefalico (bulbo, ponte e mesencefalo) e da brevi colonne effettrici viscerali site nella porzione sacrale del MS da S2 a S4. E’ da sottolineare che ogni viscere riceve una doppia innervazione vegetativa, cioè fibre postgangliari parasimpatiche e ortosimpatiche, ad azione antagonista. Altre differenze riguardano la collocazione dei gangli e i mediatori chimici utilizzati. Nell’ortosimpatico per esempio, i gangli sono vicino al centro nevrassiale e ciò determina la presenza di fibre pregangliari molto corte e postgangliari molto lunghe. Nel parasimpatico invece, i gangli sono posti addirittura nello spessore del viscere innervato ma ci posso essere molte eccezioni. Per quanto riguarda neuro trasmettitori, è logico che essendo le funzioni di ciascuna innervatura antagoniste, il mediatore chimico usato sarà diverso per un’azione eccitatoria o inibitoria. Il sistema nervoso ortosimpatico (simpatico) è composto da un'unità centrale di controllo, l'ipotalamo, e da una serie di coppie di neuroni efferenti (che emettono segnali in uscita). Il primo neurone (pregangliare), localizzato nel corno laterale dei segmenti spinali che vanno dalla prima vertebra toracica (T1) alla seconda lombare (L2), emette un assone che si diparte, attraverso la radice anteriore del midollo spinale, insieme al nervo spinale, da cui però si divide appena fuori dal foro di coniugazione spinale (foro intervertebrale) dirigendosi, come breve ramo comunicante bianco (fibre pregangliari mieliniche), al ganglio della catena dell'ortosimpatico del segmento spinale corrispondente (fa eccezione a questa regola l'innervazione della midollare del surrene). Le catene dell'ortosimpatico appaiono come due fili di perle poste parallelamente, una destra e l'altra a sinistra, lungo tutta la colonna vertertebrale. I suoi gangli (le "perle") sono connessi fra di loro da sottili fasci nervosi (ciò consente in realtà a una fibra pregangliare di prendere contatti anche con altri segmenti, sia superiori che inferiori). Da qui i segnali, direttamente o passando da ulteriori gangli, arrivano, tramite le fibre postgangliari (amieliniche), ai vari organi bersaglio. Le fibre pregangliari ortosimpatiche utilizzano come neurotrasmettitore l'acetilcolina e il peptide vasointestinale (VIP), quelle postgangliari la noradrenalina, l'adrenalina, la dopamina e il neuropeptide Y (NPY) . Anche il sistema nervoso parasimpatico è regolato dall'ipotalamo. Anatomicamente l'innervazione efferente parasimpatica decorre lungo i nervi cranici, di cui il X o nervo vago innerva gli organi della cavità toracica e addominale (quasi esclusivamente dipendenti da questo nervo) e i restanti raggiungono gli organi cranici, e lungo gli assoni dei neuroni parasimpatici presenti nei corni laterali dei segmenti spinali che vanno dalla seconda alla quarta vertebra sacrale (S2-S4). Questi ultimi decorrono poi quasi tutti nel nervo pudendo innervando organi genitali, vescica e intestino retto. Tutti i nervi parasimpatici contengono esclusivamente fibre pregangliari (che utilizzano come neurotrasmettitori l'acetilcolina e il peptide vasointestinale VIP) in quanto, a differenza del sistema nervoso ortosimpatico, i gangli parasimpatici sono situati in prossimità o all'interno degli organi innervati. La maggior parte degli organi è innervata da entrambe le componenti (ortosimpatica e parasimpatica). Vi sono però alcune parti del corpo che ricevono un’innervazione esclusivamente simpatica. Tra queste, la più importante è la porzione midollare delle surrenali (due piccole ma importantissime ghiandole endocrine poste ognuna sopra il polo superiore dei reni). Inoltre, la midollare del surrene viene innervata, attraveso i nervi splancnici, da fibre nervose pregangliari del simpatico che non si interrompono, ovvero che non formano sinapsi con i gangli della catena paravertebrale dell'ortosimpatico, ma che si connettono direttamente a un determinato gruppo di cellule della midollare, cellule cromaffini (formando in sostanza un ganglio interno), stimolandole a produrre le catecolamine adrenalina, noradrenalina e, in minima quantità, dopamina. Ci troviamo qui di fronte a una via direttissima che collega il cervello, tramite il sistema nervoso simpatico, alle surrenali. La visione del pericolo, infatti, mette in funzione questa via immediata, inducendo a produrre, da parte delle cellule della midollare del surrene, una miscela “allarmante” composta per l’80% di adrenalina e il 20% di noradrenalina, in quantità dieci volte maggiore del normale. Se però il pericolo perdura, allora entreranno in scena ulteriori significativi cambiamenti che coinvolgeranno ancora una volta le surrenali (questa volta la zona corticale), organi primari della reazione di stress. Gli effetti del sistema nervoso ortosimpatico sono facilmente comprensibili se interpretati in relazione alla "reazione di attacco o difesa" (stress fisico e psichico). In molti casi il sistema nervoso parasimpatico è suo antagonista. Inoltre il sistema nervoso vegetativo invia al midollo spinale sensazioni quali, per esempio, dolore, stato di riempimento dello stomaco e della vescica. Tali fibre afferenti arrivano normalmente alla radice posteriore del midollo spinale non distinguendosi però in ortosimpatiche o parasimpatiche. Pertanto, poichè nei segmenti del midollo spinale convergono afferenze della sensibilità dolorifica degli organi interni e afferenze cutanee, il dolore viscerale puà essere proiettato e quindi percepito sulle aree cutanee, dermatomeri, del relativo segmento midollare (cuore-faccia interno braccio sinistro, cistifellea-addome superiore destro, pancreasaddome sinistro ecc.). Descrivendo il sistema nervoso autonomo occorre ricordare l'esistenza di una terza componente vegetativa: il sistema nervoso metasimpatico, importnate quanto le altre due (ortosimpatica e parasimpatica). Il sistema nervoso metasimpatico è presente all'interno delle pareti dell'intestino e dello stomaco come rete nervosa formata da circa cento milioni di neuroni, collegata alle fibre nervose ortsimpatiche e parasimpatiche (plesso sottomucoso di Meissner e motorio mienterico di Auerbach). Ricerche recenti hanno potuto affermare che questa rete svolge un ruolo in gran parte indipendente dal sistema nervoso centrale (SNC). Data l'importanza attruibuita dai fisiologi a tale struttura, essa viene definita come "cervello addominale o "cervello enterico". E' chiaro, data la grande attiguità, che l'alimentazione avrà sul sistema metasimpatico una grossa influenza. In conclusione va detto che la classica netta separazione tra il sistema nervoso vegetativo, considerato come controllore delle funzioni viscerali in maniera indipendente dalla nostra volontà, e il sistema nervoso periferico volontario, è oggi sempre più messa in discussione da qualificati studiosi, in special modo dopo l'avvento della psiconeuroimmunologia. Il sistema nervoso costituisce una fittissima rete di comunicazione (solo nel cervello vi sono 10 miliardi di neuroni) diffusa in tutto il corpo e strettamente interconnessa con due altri importanti sistemi di regolazione dell'organismo: il sistema endocrino e il sistema immunitario. I nervi cranici o encefalici I nervi cranici, o nervi encefalici, sono un gruppo di nervi che invece di avere origine dal midollo spinale, partono direttamente dal tronco encefalico. Nell'anatomia umana ci sono dodici paia di nervi cranici pari (destri e sinistri), numerati dall'alto verso il basso con numeri romani: 1. nervo olfattivo (I) Hanno le stesse caratteristiche dei nervi spinali in quanto presentano fibre motrici e fibre 2. nervo ottico (II) sensitive, viscerali e somatiche. La differenza sta nel fatto che i nervi encefalici possono 3. nervo oculomotore (III) avere anche solo una di queste caratteristiche, mentre i nervi spinali sono nervi misti. Questa 4. nervo trocleare (IV) è la classificazione convenzionale dei nervi cranici; tuttavia alcuni di questi hanno 5. nervo trigemino (V) caratteristiche del tutto particolari che non si riscontrano altrove: per esempio il nervo ottico e 6. nervo abducente (VI) il nervo olfattivo sono delle estensioni del cervello e quindi a rigor di termini non potrebbero 7. nervo faciale (VII) essere considerati dei veri nervi. Il nervo faciale è costituito da due nervi distinti: nervo 8. nervo vestibolococleare (VIII) faciale propriamente detto e nervo intermedio del Wrisberg. Il nervo vestibolo-cocleare o 9. nervo glossofaringeo (IX) stato-acustico consta di due nervi distinti: nervo cocleare e nervo vestibolare. Il nervo 10. nervo vago (X) accessorio consta dell'unione di due nervi: nervo accessorio del vago e nervo accessorio 11. nervo accessorio (XI) spinale; però bisogna considerare che il nervo accessorio spinale non ha origine dal cranio, 12. nervo ipoglosso (XII) ma si estende dal V segmento cervicale del midollo spinale verso l'alto, fino all'estremità inferiore del midollo allungato ed entra nella cavità del cranio attraverso il foro occipitale, dove si unisce al nervo accessorio del vago. Quindi, data la sua origine extra-cranica il nervo accessorio spinale non potrebbe essere considerato propriamente un nervo cranico. Fatte queste premesse e soprattutto se ritenute veritiere e quindi valide, la classificazione convenzionalmente accettata dei nervi cranici potrebbbe essere modificata. Quasi tutti i nervi cranici sono connessi a dei nuclei di materia grigia all'interno del tronco encefalico, e gli assoni da e per i nervi cranici incontrano le loro sinapsi all'interno di questi nuclei. Danneggiare uno di questi nuclei associati ha lo stesso effetto di una resezione (taglio) del nervo cranico corrispondente, con conseguente cecità dal lato ipsilaterale. Come nel midollo spinale, anche i nuclei del tronco encefalico sono divisi in ventrali (anteriori) e dorsali (posteriori), e nello stesso modo i nuclei ventrali sono somatici, cioè controllano muscoli scheletrici, mentre i nuclei dorsali sono sensoriali. Vicino al piano di simmetria del tronco encefalico ci sono i nuclei efferenti somatici, come i nuclei oculomotori, mentre lateralmente ci sono i nuclei efferenti autonomi (viscerali) come il nucleo di Edinger-Westphal, che controlla le lacrime. Procedendo verso l'esterno si incontra una separazione, detta sulcus limitans, oltre la quale troviamo i nuclei viscerali afferenti, come il nucleo del solitario; più lateralmente ancora, ma più in avanti, troviamo troviamo il nucleo trigemino e dorsalmente al tronco gli afferenti somatici specifici, che gestiscono sensazioni come l'equilibrio. I nervi spinali nervi che nascono dal midollo spinale con radici dorsali e ventrali; le prime sono sensorie e ospitano nel loro decorso un ganglio; le radici ventrali sono motrici e sprovviste di gangli, si uniscono a formare i nervi spinali misti. Il nervo misto si divide poi in un ramo dorsale e in uno ventrale; quest’ultimo possiede 4 tipi di fibre: somatomotrici e visceromotrici, somatosensitive e viscerosensitive. Il ramo dorsale non ha fibre visceromotrici. Dal ramo ventrale parte poi una diramazione, o ramo comunicante, che termina in un ganglio simpatico, per cui il sistema nervoso simpatico è controllato dal sistema nervoso centrale. Le 31 paia di nervi spinali si suddividono in: 8 cervicali, 12 toracici, 5 lombari, 5 sacrali e 1 paio di nervi coccigei. Formano vari plessi: plesso cervicale, formato dai primi 4 nervi cervicali; plesso brachiale, formato dagli altri nervi cervicali e dal primo toracico (gli altri nervi toracici non formano plessi); plesso lombare, formato dai primi 4 nervi lombari; plesso sacrale, formato dall’ultimo nervo lombare e dai primi 4 sacrali. L’ultimo nervo sacrale e il coccigeo formano il plesso sacro-coccigeo. È inoltre da ricordare che i nervi per gli arti derivano dai plessi cervico-brachiale e lombo-sacrale; quest’ultimo tende a separarsi in due plessi distinti, lombare e sacrale; dal plesso lombare si originano alcuni nervi, fra cui l’otturatore e il femorale, e dal sacrale il nervo ischiatico. PARTI DEL SISTEMA NERVOSO TRONCO DEL CORPO CALLOSO COLONNA DEL FORNICE III VENTRICOLO PLESSO CORIOIDEO DEL III VENTRICOLO ARTERIA CEREBRALE ANTERIORE COMMESSURA ANTERIORE CHIASMA OTTICO IPOTALAMO ARTERIA CEREBRALE POSTERIORE IPOFISI ARTERIA BASILARE CERVELLETTO ARTERIA VERTEBRALE PONTE BULBO IV VENTRICOLO CANALE EPENDIMALE
Scaricare