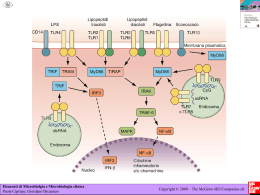
Rivista Italiana di Genetica e Immunologia Pediatrica - Italian Journal of Genetic and Pediatric Immunology Anno I numero 3 - ottobre 2009 | direttore scientifico: Carmelo Salpietro - direttore responsabile: Giuseppe Micali Infezioni virali e circuiti immunologici Viral infections and immunological circuits Caterina Cuppari, Piera Vicchio, Silvana Briuglia, Romina Gallizzi, Carmelo Salpietro Dipartimento di Scienze Pediatriche, UOC Genetica e Immunologia Pediatrica, Università di Messina Abstract In recent years have greatly increased our knowledge of how viruses are recognized by the innate immune system and the functional consequences of 'virus-host interaction. The innate immune system consists of a variety of cellular effectors, directly or through their products are capable of inducing an immune response that, if sufficient, it prevents the spread of the pathogen and the onset of the disease. In the early stages of infection and then activate a circuit of immunoregulatory cytokines communication between various cell populations was the result of lead, strengthen and support the coordinated activity of different populations of the innate response, and to condition the environment cytokine responsible for investigation of the adaptive response. The knowledge of the mechanisms of virus-immune cells leads to increased understanding of why in the same community or in the same family some individuals exposed to the same risk of infection have a clinical expression of disease as well as differentiated and closely correlated with the virulence of the germ, with more or less complete integrity of the entire molecular network defense. Similarly, the depth of interactive circuits virus-immune system opens up new prospects for prevention. Knowing the close link between the innate and adaptive immunity the implications are relevant for the design of vaccine strategies and immunoterapeutiche. Other knowledge will allow more than a specific preventive and therapeutic strategies too early to acquire information in order to counter also the variability viral antigens. Riassunto Negli ultimi anni sono notevolmente aumentate le nostre conoscenze su come i virus vengono riconosciuti da parte del sistema immunitario innato e le conseguenze funzionali dell’interazione virus-ospite. Il sistema immunitario innato è costituito da una varietà d'effettori cellulari che direttamente o attraverso prodotti sono in grado di indurre una risposta immunitaria che, se sufficiente, previene la diffusione del patogeno e l'insorgenza della malattia. L'immunità naturale è inoltre in grado di regolare l'attivazione del sistema immunitario specifico, e quindi la produzione di anticorpi (immunità umorale) e cellule T antigene-specifiche (immunità cellulare). Discussione L'immunità naturale innata si mette in atto entro pochi minuti dall'aggressione ed è mediata fondamentalmente dalle cellule fagocitarie (monociti/macrofagi, le cellule dendritiche e i polimorfonucleati, dalle cellule di citotossicità naturale NK) e da prodotti solubili, quali gli interferoni (IFN), le citochine, le chemochine. Le cellule dendritiche rappresentano un ponte tra il sistema immune naturale ed antigene specifico, ed esse, come tutte le cellule dell'immunità naturale, esprimono recettori (Pathogen Recognition Receptors: PRRs) in grado di riconoscere strutture caratteristiche di gruppi di microrganismi (pathogen associated molecular patterns: PAMPs). I "Toll-like receptors" (TLR) rappresentano al momento una delle classi più studiate di PRR. Utilizzando specifiche combinazione di TLRs le cellule dell'immunità naturale sono capaci di discriminare tra le varie classi di patogeni e, quindi, di reagire attivando direttamente una risposta in grado di contenere la diffusione del patogeno oppure stimolando e/o indirizzando nel giusto verso la risposta immunitaria acquisita. L’ attivazione virale dei TLR catalizza una complessa cascata di segnali, non ancora completamente chiariti, che inizia dal dominio citoplasmatico del recettore toll/interleukin-1 (TIR), che lo condurrà all’ espressione di diversi geni portando alla rapida produzione di citochine proinfiammatorie, alla maturazione delle cellule dendritiche e favorendo una risposta orientata verso un profilo di tipo T helper 1. In particolare i TIR domain contengono molecole adattatrici, come MyD88, (utilizzata da tutti i TLRs eccetto il TLR3), TIRAP, TRIF, e TRAM (per TLR4), che portano alla reclutazione dei complessi IRAKs e TRAFs attraverso il segnale NFkB14. In ultima analisi, la stimolazione TLR culmina nella sintesi di citochine antivirali, come IFN, IL-1β e IL-6, che possono direttamente sopprimere la replicazione virale (fig.1). Fig. (1) TLR signal transduction pathway Da: Toll-Like Receptors and Viruses: Induction of Innate Antiviral Immune Responses Angeliki Xagorari1 and Katerina Chlichlia The Open Microbiology Journal, 2008, 2, 49-59 Questi segnali reclutano le cellule immunitarie ai siti di infezione virale e attivano le cellule dendritiche (DC). Le cellule esprimono anche delle RNA elicasi citoplasmatiche che funzionano come una classe di recettori attraverso il riconoscimento dell’ RNA a doppio filamento (dsRNA) prodotto durante la replicazione del virus. Queste due classi di molecole recettoriali di riconoscimento, i TLR e RNA-elicasi, espressi in diversi compartimenti intracellulari, inducono una risposta mediata dell’interferon di tipo I attraverso vie distinte di segnale. I virus sono caratterizzati dalla complessità dei loro genomi e sono classificati in base al loro meccanismo di replicazione (classificazione Baltimore). Gli acidi nucleici del genoma virale possono essere DNA o RNA, a polarità positiva o negativa, a singolo o doppio filamento, a molecola continua (a volte circolare) o con configurazione segmentata. Quattro tipi di TLR sembrano giocare un ruolo critico nel riconoscimento degli acidi nucleici dei virus: 3, 7, 8 e 9. Tutti e quattro i recettori riconoscono acidi nucleici e per la loro attivazione è necessaria l'acidificazione endosomiale. Il TLR3 riconosce dsRNA (dsRNA costituisce il genoma di una classe di virus, ma è anche generato durante il ciclo vitale di molti virus), TLR7 e 8 riconoscono il singolo filamento di RNA (ssRNA) e TLR9 risponde a dsDNA riconoscendo le porzioni non-metilate CpG che contengono DNA virale. Altri TLR sono coinvolti nel riconoscimento virale; TLR2 e TLR4 riconoscono per esempio glicoproteine del rivestimento virale. Nelle fasi precoci di infezioni quindi l’attivazione immunoregolatoria di un circuito citochinico di comunicazione tra le varie popolazioni cellulari ha il risultato di indurre, potenziare e sostenere l'attivazione coordinata delle diverse popolazioni della risposta innata, e di condizionare l'ambiente citochinico responsabile dell'istruzione della risposta adattativa. Dati recenti suggeriscono che mutazioni o variazioni funzionali (polimorfismo) a carico dei recettori Toll-like, o delle proteine che partecipano alla trasduzione del segnale intracellulare, o dei geni che codificano per varie citochine pro- e anti-infiammatorie possono associarsi ad una aumentata suscettibilità verso particolari agenti infettivi o rendere più grave il decorso della malattia. Questo è quello che accade per esempio nella sindrome di Duncan, rara malattia X-linked, dove l’infezione da Virus di Epstein-Barr (EBV) risulta essere fatale. In un soggetto normale l’infezione da EBV di cellule B provoca l'attivazione delle cellule B e la loro proliferazione. La via di segnale dei TLR fornisce un terzo dello stimolo di attivazione delle cellule B. L’EBV up-regola l'espressione di TLR-7 e down-regola l'espressione di TLR-9 in cellule B naïve. IRF-5 è indotto in seguito ad infezione da EBV e la proliferazione delle cellule B fornisce l’evidenza che EBV modula il segnale del TLR-7. Dopo l'infezione da EBV è indotta la trascrizione di una nuova variante splice di IRF-5 ed agisce come una forma dominante negativa. L’EBV quindi utilizza inizialmente il segnale del TLR-7 per migliorare la proliferazione cellulare B e successivamente modifica la via per l’attività regolatoria di IRF-5. E’ stato dimostrato che anche il TLR2 svolge un’ importante ruolo nella risposta immunitaria nei confronti dell’ infezione da EBV. Virioni di EBV infettivi hanno dimostrato di portare alla attivazione di NF-B attraverso iTLR-2. Inoltre, l'infezione primaria da EBV in monociti umani induce il rilascio di una chemochina, la proteina 1 chemiotattica per i monociti (MCP-1), e piccoli RNA che interferiscono con i TLR2 per ridurre in modo significativo la risposta di chemochine contro EBV. La proliferazione dei linfociti B infettati dal virus è controllata in condizioni normali dall’ attivazione dei linfociti T da parte delle citochine pro-infiammatorie (IL-6, IL-12). Questo non accade nella Sindrome di Duncan (o sindrome linfoproliferativa), dove l’infezione da EBV si dimostra estremamente grave potendo portare direttamente alla morte oppure evolvere in un linfoma, in un’aplasia midollare oppure in una ipogammaglobulinemia. È stato ipotizzato che i soggetti colpiti, immunologicamente normali fin al momento dell’infezione da virus Epstein Barr presentino una risposta immunitaria, mediata da un linfocita T aberrante (controllato da un gene presente sul cromosoma X), che non è in grado di controllare la proliferazione dei linfociti B, con conseguente distruzione delle cellule linfoidi ed epatiche del paziente. In particolare è dovuta ad una mutazione a carico della proteina associata (PSA) alla molecola di segnalazione di attivazione linfocitaria (SLAM) (fig2). Nella tabella 1 sono riportati nello specifico i vari tipi di virus e i rispettivi TLR interessati in particolare. ll Citomegalovirus (CMV) è un virus dsDNA che innesca l'attivazione di TLR durante il contatto e/o l’ingresso virus-cellula. Il TLR2 riconosce le glicoproteine dell’envelope CMV B e H, portando a una risposta immunitaria innata da attivazione di NF-kB e l'induzione di citochine infiammatorie. Il riconoscimento del CMV è mediato anche dall’ TLR9 e dal TLR3 con la produzione di interferone di tipo I. per avere una protezione completa è necessario che tutti e tre recettori agiscano insieme. L’ Herpes simplex virus di tipo 1 (HSV1) è un dsDNA virus ed è la causa principale di sporadiche encefaliti letali e cecità. L’HSV1 induce la morte cellulare per apoptosi delle cellule della microglia. Oltre al ruolo di TLR2 in caso di infezioni da CMV, il TLR2 è attivato in risposta al virus della varicella Zoster (VZV) e sembra svolgere un ruolo significativo nelle infezioni da HSV1. Sembra che la resistenza innata a HSV1 è mediata da MyD88 e può essere attivata da TLR multipli, in particolare dapprima si attiva l’ TLR3 con la produzione di l’ IL6 e IRF1 e ed in una seconda fase si ha l’attivazione del TLR7 mediata da NF-kB. Le sequenze CpG, molto presenti nel genoma di HSV1 e HSV2 sono riconosciuti da TLR9 espresso soprattutto sulle cellule dendritiche polmonari. L’Adenovirus (ADV) è un ds DNA virus che causa malattie respiratorie, oculare e gastrointestinali. L’ADV attiva l’immunità innata attraverso vie TLR dipendentI e indipendenti. Il riconoscimento dell’ADV da parte delle DC polmonari è mediato dal TLR9 e dipende da MyD88 che svolge un ruolo significativo come un amplificatore e regolatore di immunità adenovirali. Al contrario, il riconoscimento dell’ADV da parte di cellule DC non plomonari è TLR-indipendente ed avviene attraverso il rilevamento citosolico del DNA. Il Papillomavirus umano (HPV) è un altro virus dsDNA; molti dati suggeriscono che l'infezione da HPV è causa del cancro della cervice uterina. L'infezione da virus HPV 16L1 - come particelle virali (VLPs), avvia l'immunità per attivazione delle DC e una risposta potente di IgG neutralizzanti, che richiede la via di segnale MyD88dipendente. IFN-alpha/beta così come numerose citochine proinfiammatorie e chemochine sono upregulated in risposta ad una infezione da HPV16 VLPs. Cellule DC derivate dal midollo osseo e carenti di MyD88 falliscono la up-regolazione dell’IL12, IFN-alfa e IFN-gamma in risposta a HPV16VLPs. Inoltre, risposte immunitarie Th1-mediate sono ridotte nei topi deficienti in MyD88 e in recettori per IFN-alpha/beta. Queste osservazioni implicano che i TLR abbiano un ruolo centrale nel riconoscimento immunitario di VLPs HPV16L1. L'internalizzazione del retrovirus dell'immunodeficienza umana virus di tipo 1 (HIV-1) è mediata da interazioni dell’involucro virale con il recettore CD4 dell’ospite. Gli RNA virali presenti nei compartimenti endosomiali e riconosciuti da TLR7 inducono la secrezione di IFN-alfa. Gli stessi RNA ricchi di Uridina dell’ HIV-1 sono potenti attivatori delle cellule NK attraverso i TLR7 / 8. L’associazione costitutiva di MyD88 con IRAK1 è evidente nel virus di tipo 1 della leucemia (HTLV-1)- cellule T trasformate; HTLV-1 Tax aumenta l'espressione di TLR e attiva sinergicamente NF-kB attraverso il MyD88 wt. Gli HTLV-1 hanno anche sviluppato una proteina che interferisce con la via di segnale del TLR4. HTLV-1 p30 interagisce con PU.1 e inibisce la sua attività di legame al DNA e la trascrizione conseguente con down-regolazione dell'espressione del TLR4 dalla superficie cellulare]. Espressione di p30 ostacola il rilascio di citochine proinfiammatorie e stimola il rilascio di citochine antinfiammatorie come IL-10 in seguito a stimolazione di TLR4 nei macrofagi umani. Questa nuova funzione di p30 può spiegare la ridotta attivazione dell’immunità adattativa in pazienti adulti con cellule T- leucemiche (ATL). TLR4 induce anche la maturazione delle cellule dendritiche derivate dal midollo osseo e l'up-regolazione del recettore d’entrata MMTV (CD71), su queste cellule. Il Virus dell'epatite C (HCV), virus ssRNA, è in grado di determinare infezioni croniche. Patogeni sono comuni in pazienti con infezione da HCV e possono essere riconosciuto da TLR, che sono upregulated in monociti e cellule T. Le DC riconoscono l’Infezioni da virus attraverso la via dei TLR e RIG e producono grandi quantità di IFN di tipo I e citochine infiammatorie. La ridotta espressione di TLR2 su DC immature di pazienti con infezione da HCV rispetto al controllo dimostrano una minore capacità di stimolare la proliferazione delle cellule T. Il Virus dell'epatite C purificato da siero dei pazienti con infezione cronica in forma di lipoviro - particelle (LVP, delle lipoproteine ricche in trigliceridi come particelle contenente l'RNA virale e proteine) interagisce con TLR4 e attiva la maturazione di DC e di induce attraverso meccanismi ERK e P38- dipendenti un fenotipo Th2, invece di un Th1. Cellule Th2 producono IL4, IL5, IL6, IL9, IL10 Il13 e citochine che promuovono l'espansione delle cellule B e downregulano le Cellule Th1. Plasmacitoidi DCS (PDC), sono note per secernere grandi quantità di IFN-alfa in risposta all'attivazione TLR e sono in grado di attivare le cellule T naïve. In pazienti con infezione da HCV le PDC possono mostrare una ridotta capacità di risposta al legame con TLR (R848 stimolazione), accompagnata da ridotta espressione dei marker HLA-DR e di citochine IFN-alfa, e da una compromessa attivazione di cellule T-CD4+ naive. In DC mieloidi di pazienti con infezione da HCV, i livelli di IFN-beta mediati da TLR / RIG o l’ induzione di TNF-alfa sono inferiori rispetto a quelle dei donatori non infetti, nonostante la più alta espressione di TLR2, TLR4 e RIG-1, il che significa che la trasduzione del segnale è ridotto in HCV-cellule infette. Ci sono molti studi che dimostrano che componenti di HCV possono legarsi a TLR e attivare la loro via di segnale o bloccarne la funzione interferendo con intermediari intracellulari. Altri studi hanno dimostrato un ruolo immunomodulante della proteina core HCV. La Proteina core HCV può associarsi con il recettore della superficie cellulare del componente del riconoscimento C1, recettore gC1q (gC1qR) su DC umani e inibire la produzione di IL12 TLR4-indotta senza compromettere la produzione di altre citochine TLR-indotte. Allo stesso modo, l'incubazione di cellule mononucleate con proteine del core HCV determinano un upregulation dell’espressione dei TLR2 e la soppressione di TLR4 e TLR7 in pazienti e controlli. I TLR2 possono utilizzare TLR1 e TLR6 come corecettori per l'attivazione dei TLR2 dei macrofagi e dell’immunità innata nell'uomo e nel topo. Studi con linee cellulari di macrofagi del topo esprimendo stabilmente le proteine non strutturali HCV (NS3, NS3/4A, NS4B o NS5A) hanno dimostrato l'inibizione delle vie di segnale di TLR2, TLR4, TLR7 e TLR9. La soppressione dell’espressione del TLR3 può essere responsabile della persistenza del virus in caso di infezione cronica da HCV. Il Virus dell'epatite B (HBV) è un virus dsDNA, che può causare infezioni acute e croniche. L'antigene di superficie del virus dell'epatite B (HBsAg) è più frequentemente usato per individuare la presenza di questa infezione virale. Diversi TLR sono coinvolti nel controllo dell’infezione HBV. Iniezione endovenosa del virus in topi transgenici TLR2, TLR3, TLR4, TLR5, TLR7 e TLR9 ha dimostrato che tutti i leganti, tranne TLR2 inibiscono la replicazione del virus dell'epatite B nel fegato senza causare danno cellulare entro 24 ore in un modo IFNalpha / beta-dipendente. Altri studi riportano che il TLR2 è ridotto nelle cellule mononucleate del sangue periferico infettate da HBV, mentre è più alta l’espressione TLR4 rispetto agli altri TLR. L’intima conoscenza dei meccanismi di interazione virus-cellule immunitarie comporta una sempre maggiore comprensione del perché in una stessa comunità o in una stessa famiglia alcuni soggetti esposti allo stesso rischio infettivo hanno un’espressività clinica di malattia differenziata e strettamente correlata oltre che con la virulenza del germe, anche con l’integrità più o meno completa di tutto il network molecolare di difesa. Nostre ricerche in corso incominciano ad evidenziare per esempio un particolare identikit genetico-immunologico di alcuni pazienti che hanno avuto una infezione da EBV clinicamente importante. Analogamente l’approfondimento di circuiti interattivi virus-sistema immunitario apre la strada a nuove prospettive di prevenzione. Conoscere lo stretto legame esistente fra l'immunità innata e quella adattativa ha infatti delle implicazioni rilevanti per la progettazione di strategie vaccinali ed immunoterapeutiche. Per esempio, per la loro capacità di attivare potentemente le cellule dell'immunità innata, è stato recentemente proposto il possibile utilizzo dei ligandi dei TLR come adiuvanti vaccinali in strategie terapeutiche di immunostimolazione (fig.3). La cooperazione fra i diversi effettori dell'immunità naturale (eventualmente potenziata dall'uso immunoterapeutico dei ligandi dei TLR) potrebbe essere anche efficace nel superare le strategie di evasione dei patogeni. I TLR agonisti rappresentano quindi un approccio promettente per il trattamento delle malattie infettive. Una conoscenza più approfondita dei circuiti regolatori dell'immunità innata ed il loro impatto sui partecipanti alla risposta adattativa è quindi necessaria per una progettazione razionale di nuove strategie immunomodulatrici. Ulteriori conoscenze potranno consentire oltre una specifica strategia preventiva e terapeutica anche di acquisire informazioni precocemente al fine di contrastare anche la variabilità antigenica virale. Bibliografia • Kawai T, Akira S. Pathogen recognition with Toll-like receptors. Curr Opin Immunol 2005; 17: 338-44 • Kawai T, Akira S. Innate immune recognition of viral infection. Nat Immunol 2006; 7 (2): 131-37. • Pasare C, Medzhitov R. Toll-like receptors: linking innate and adaptive immunity. Microbes Infect 2004; 6 (15): 1382-87. • Meylan E, Tschopp J. Toll-like receptors and RNA helicases: two parallel ways to trigger antiviral responses. Mol Cell 2006; 22 (5): 561-9. • Bowie A, Kiss-Toth E, Symons JA, Smith GL, Dower SK, O’Neill LA. A46R and A52R from vaccinia virus are antagonists of host IL-1 and toll-like receptor signalling. Proc Natl Acad Sci USA 2000; 97: 10162-167. • Sato A, Iwasaki A. From The Cover: Induction of antiviral immunity requires Toll-like receptor signaling in both stromal and dendritic cell compartments. Proceedings of the National Academy • of Sciences 2004;101:16274–16279. • Stack J, et al. Vaccinia virus protein A46R targets multiple Toll-like-interleukin-1 receptor adaptors and contributes to virulence. J Exp Med 2005;201:1007–1018. [PubMed: 15767367] • Akira S. TLR signaling. Curr Top Microbiol Immunol 2006;311:1–16. [PubMed: 17048703] • Uematsu S, Akira S. Toll-like receptors and innate immunity. Journal of Molecular Medicine 2006;84:712–725. • Marco C. TLR pathways and IFN-regulatory factors: To each its own. European Journal of Immunology 2007;37:306–309. [PubMed: 17273997] • Martin HJ, Lee JM, Walls D, Hayward SD. Manipulation of the Toll-like receptor 7 signaling pathway by Epstein-Barr virus. J Virol 2007; 81 (18): 9748-58. • Toll-Like Receptors and Viruses: Induction of Innate Antiviral Immune Responses Angeliki Xagorari1 and Katerina Chlichlia The Open Microbiology Journal, 2008, 2, 49-59 • Boehme KW, Guerrero M, Compton T. Human cytomegalovirus envelope glycoproteins B and H are necessary for TLR2 activation in permissive cells. J Immunol 2006; 177: 7094-102. • Compton T, Kurt-Jones EA, Boehme KW, et al. Human cytomegalovirus activates inflammatory cytokine responses via CD14 and Toll-like receptor 2. J Virol 2003; 77 (8): 4588-96. • Tabeta K, Georgel P, Janssen E, et al. Toll-like receptors 9 and 3 as essential components of innate immune defense against mouse cytomegalovirus infection. Proc Natl Acad Sci USA 2004; 101 (10): 3516-21. • Lore K, Betts MR, Brenchley JM, et al. Toll-like receptor ligands modulate dendritic cells to augment cytomegalovirus- and HIV-1- specific T cell responses. J Immunol 2003; 171 (8): 4320-8. • Lund J, Sato A, Akira S, Medzhitov R, Iwasaki A. Toll-like receptor 9-mediated recognition of Herpes simplex virus-2 by plasmacytoid dendritic cells. J Exp Med 2003; 198 (3): 513-20. • Ashkar AA, Bauer S, Mitchell WJ, Vieira J, Rosenthal KL. Local delivery of CpG oligodeoxynucleotides induces rapid changes in the genital mucosa and inhibits replication, but not entry, of herpes • simplex virus type 2. J Virol 2003; 77 (16): 8948-56. • Zhu J, Huang X, Yang Y. Innate immune response to adenoviral vectors is mediated by both Toll-like receptor-dependent and – independent pathways. J Virol 2007; 81 (7): 3170-80. • Hartman ZC, Kiang A, Everett RS, et al. Adenovirus infection triggers a rapid, MyD88-regulated transcriptome response critical to acute-phase and adaptive immune responses in vivo. J Virol2007; 81 (4): 1796-812. • Dolganiuc A, Garcia C, Kodys K, Szabo G. Distinct Toll-like receptor expression in monocytes and T cells in chronic HCV infection. World J Gastroenterol 2006; 12 (8): 1198-204. • Sato K, Ishikawa T, Okumura A, et al . Expression of Toll-like receptors in chronic hepatitis C virus infection. Gastroenterol Hepatol2007; 22 (10): 1627-32 • Mazzoni A, Segal DM. Controlling the Toll road to dendritic cell polarization. J Leukoc Biol 2004;75 (5): 721-30. • Yakushijin T, Kanto T, Inoue M, et al .Reduced expression and functional impairment of Toll-like receptor 2 on dendritic cells in chronic hepatitis C virus infection. Hepatol Res 2006; 34 (3): 156-62 • Agauguè S, Perrin- Cocon L, André P, Lotteau V. Hepatitis C lipoviro- particle from chronically infected patients interferes with TLR4 signaling in dendritic cell. PLoS One 2007; 2 (3): e330. • Yonkers NL, Rodriguez B, Milkovich KA, et al. TLR liganddependent activation of naïve CD4 T cells by plasmacytoid dendritic cells is impaired in hepatitis C virus indection. J Immunol2007; 187 (7): 4436-44. Trimestrale di divulgazione scientifica dell'Associazione Pediatrica di Immunologia e Genetica Legge 7 marzo 2001, n. 62 - Registro della Stampa Tribunale di Messina n. 3/09 - 11 maggio 2009 Direttore scientifico Carmelo Salpietro - Direttore responsabile Giuseppe Micali - Segreteria redazione Basilia Piraino - Piera Vicchio Direzione-Redazione: UOC Genetica e Immunologia Pediatrica - AOU Policlicnico Messina www.geneticapediatrica.it/rigip
Scaricare