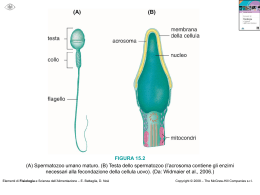
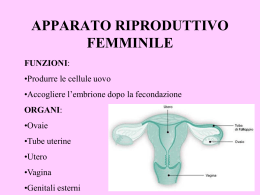
LICEO SCIENTIFICO “ARCHIMEDE” ACIREALE (CT) Prof. Alfio Francesco Cannone ANATOMIA a.s. 2007 – 2008 UMANA Anatomia Umana ATTIVITA' CELLULARE La cellula è in grado di procurarsi le sostanze alimentari, per crescere, per produrre altre molecole, per rimpiazzare parti della sua struttura e per riprodursi. Le cellule quindi hanno attività metabolica e riproduttiva autonoma, ma per crescere armonicamente hanno bisogno di comunicare fra di loro: Si procura gli alimenti Per mezzo del trasporto attivo e passivo Produce molecole ATTIVITA' Rimpiazza parti cellulari mancanti CELLULARE Cresce armonicamente RER, REL, sintesi proteica, apparato del Golgi Messaggio di crescita (fattori peptici di crescita) Generale --> G.H. Locale Si riproduce Per divisione diretta o indiretta (mitosi e meiosi) Per adesione si colloca tra cellule dello stesso tipo e si differenzia Per mezzo della matrice extracellulare --> SAM --> Somatomedine Per mezzo di molecole di riconoscimento intercellulari --> CAM Fattori peptici di crescita La crescita embrionale e quella definitiva, è regolata da messaggi di crescita che vengono inviati tramite il sanghe. Tali fattori di crescita sono: a livello generale, l'ormone G.H. (growth hormon, ormone della crescita, ormone proteico secreto dal lobo anteriore dell'ipofisi) e a livello locale le somatomedine A e B (fattori di crescita specifici per il sangue, i muscoli lisci, quelli striati….) e i fattori I° e II°. Le somatomedine, oltre che come fattori di crescita, agiscono pure sulla moltiplicazione e differenziazione cellulare. Sembra che le somatomedine funzionano da fattori di crescita perché la tirosina, presente nella molecola delle somatomedine, si fosforilizza. Ma il messaggio di crescita arriva alle cellule assieme ad altri messaggi. Ora vediamo come fa la cellula a riconoscere il G.H: Sulla membrana cellulare vi sono dei recettori di membrana (proteine intrinseche) che riconoscono solo il fattore di crescita GH e lo legano. Il complesso GH + recettore agisce su un enzima (formato da due subunità: adenilciclasi ed adenilchinasi) che si trova all'interno della membrana cellulare. La subunità catalitica (adenilciclasi ) fosforilizza l'ATP, che si trova dentro la cellula, in cAMP +P + P. La fosforilazione fa creare un poro nella membrana che serve per far entrare un flusso di (Ca++ che si lega al reticolo endoplasmatico. Il Ca++ serve per far avvenire contemporaneamente la contrazione della cellula e la trasmissione degli stimoli Il cAMP che si è formato dalla fosforilazione dell'ATP, a contatto con la subunità cinetica (adenilchinasi), fosforilizza gli istoni (proteiche basiche) legati al DNA, li despiralizza, distacca i geni, forma RNA e quindi inizia la proditosintesi, cioè l'accrescimento cellulare (si formano le proteine anche per incrementare la sostanza fondamentale dell'osso, per la crescita del muscolo……).. subunità (catalitica, adenilciclasi ) Æ ATP Æ cAMP + P + P Æ fosforilazione membrana cellulare Æ poro cellulare Æ ingresso Ca++ Æ contrazione cellulare e trasmissione stimoli subunità (cinetica, adenilchinasi) Æ cAMP Æ fosforilazione istoni Æ RNAÆ sintesi proteica Æ accrescimento cellulare Prof. A. F. Cannone 2 Anatomia Umana Interazione cellulari per mezzo della matrice extracellulare (SAM) e per mezzo di molecole di riconoscimento intercellulari (CAM) Per potersi differenziare, la maggior parte delle cellule aderisce alle cellule adiacenti per mezzo di molecole atte al riconoscimento intercellulare. Tali molecole sono le CAM, cell adhesion molecule, molecole adesive della cellula, che sono formate da glicoproteine (caderine). Oppure aderisce ad un substrato costituito da una matrice extraxellulare. Tali molecole sono le SAM (substrate adhesion molecule, molecole adesive del substrato), che sono formate da collagene, elastina, proteoglicani e glicoproteine (integrine). Tale adesione è anche una necessità, perché la cellula deve sapere dove andarsi a collocare, cioè se collocarsi in un tessuto o in un altro. Le CAM e le SAM sono delle molecole di adesione che si adattano sulle cellule e servono da proteino- -substrato su una cellula e da proteino-recettori su un’altra cellula. In tale modo le cellule se si riconoscono, formano legami stabili e rimangono legati, se invece non si riconoscono, formano legami instabili e si staccano.Quindi le CAM e le SAM, per questa caratteristica di far aderire cellule uguali fra di loro, sono indispensabili per la differenziazione cellulare e per lo sproting (la disseminazione guidata delle cellule nel sito dove devono andare a costituire un tessuto, un organo…). SAM Glicoproteine (integrine), proteoglicani, elastina, collagene CAM Glicoproteine (caderine) Meccanismi d’azione dei CAM e dei SAM Un recettore, quando arriva un ligando (CAM-SAM), produce cAMP (acido adenosinfosfato ciclico) che fosforilizza gli istoni. Ciò determina la despiralizzazione del DNA e l’inizio della proteinosintesi specifica. La sintesi proteica innesca la crescita embrionale cioè la mitosi e quindi la riproduzione cellulare. Prof. A. F. Cannone 3 Anatomia Umana APPARATO GENITALE FEMMINILE L'apparato riproduttore femminile è formato dalla vagina, dall'utero, dalle tube uterine e da 2 ovaia. Con la pubertà iniziano il ciclo ovarico, il ciclo uterino, il ciclo vaginale, che si susseguono fino alla menopausa (45-50 anni), quando la crisi nella produzione di gonadotropine ipofisarie e degli estrogeni atrofizza l'ovario ed interrompe le mestruazioni. VAGINA La vagina è un tubo muscolare lungo circa 7,5 cm, che va dalla cervice uterina, all'esterno del corpo. Si apre all'esterno, tra l'uretra (tubo proveniente dalla vescica) e l'ano. Il rivestimento interno è ricco di glicogeno che i batteri convertono in acido lattico (liquido vaginale), di conseguenza il canale vaginale ha un pH debolmente acido (4 - 5). Durante i 28 giorni del ciclo ovarico, si ha anche un ciclo vaginale. Prof. A. F. Cannone 4 Anatomia Umana UTERO L'utero è un organo muscolare cavo, a forma di pera, dalle pareti spesse. È situato fra la vescica urinaria ed il retto. Il fondo è a forma di cupola. Il corpo si trova sotto il fondo ed è separato dal collo da un leggero restringimento chiamato istmo. La cavità del corpo uterino è appiattita e di forma triangolare. Le trombe uterine si aprono nei suoi angoli basali. Il collo uterino o cervice è cilindrico, misura circa 2,5 cm di lunghezza. All'interno della cervice vi è il canale endocervicale che si apre nella vagina, nelle piccole e grande labbra e poi all'esterno, attraverso l'orifizio uterino esterno. Dal punto di vista istologico l'utero è così formato: Strato epiteliale di È formato da ghiandole (secernono il liquido uterino e muco) e da cellule rivestimento ciliate che si muovono in direzione dell'istmo dell'utero. ENDOMETRIO Lamina basale E' formata da glicoproteine e mucopèroteine. Lamina propria della È formata da tessuto connettivale in cui si trovano vasi, nervi e ghiandole che mucosa secernono muco. Per la presenza di capillari sanguigni, qui avvengono gli scambi ematici. Lamina (tonaca) È formata da connettivo denso. È la struttura portante dell'utero. sottomucosa MIOMETRIO Lamina muscolare È formata da tessuto muscolare liscio (involontario) molto spesso per permettere il travaglio del parto. PARAMETRIO È formato da linfonodi, nervi, vasi che poi arrivano nel miometrio. Il peritoneo copre il fondo ed il corpo dell'utero sulla faccia anteriore e posteriore. Il peritoneo è formato da due foglietti: esterno (a contatto con la parete addominale), interno (a contatto con gli organi dell'addome e a contatto con tutto l'utero anteriormente e posteriormente). Dall'unione del peritoneo interno anteriore e posteriore si forma il legamento largo dell'utero. Prof. A. F. Cannone 5 Anatomia Umana TUBE UTERINE Le trombe uterine o di Falloppio o ovidotti, sono due canali muscolari a forma di tromba, che si estendono dagli angoli superiori del fondo uterino (entro i margini liberi dei legamenti larghi dell'utero), fino alle ovaie. Sono costituite da tre segmenti: segmento interstiziale che arriva nell'utero, istmo, parte di mezzo, ed ampolla che termina nell'infundibolo che è sfrangiato e forma le fimbrie. Nelle fimbrie vi sono cellule secernenti ciliate. Sia l'infundibolo fimbriato che l'ampolla della tuba, non sono circondati dal legamento largo. Fra la tromba uterina e l'ovaio si forma un legamento detto mesosalpinge. OVAIA Le ovaia sono corpi grigio-rosa, solidi, di grandezza all'incirca di una mandorla (2,5 - 5 cm), situate da ciascun lato dell'utero, dietro e sotto le trombe uterine. Sono ricoperti da un mesotelio di origine epiteliale celomatico. Sotto vi è una zona midollare (ricca di vasi linfatici, sanguigni, nervi) ed una zona corticale (periferica in cui vi sono immersi i follicoli oofori in vari stati di maturazione, che contengono gli ovociti che si sono formati per ovogenesi e le cellule interstiziali. I follicoli oofori primari hanno forma sferica e sono circondati da una parete di cellule epiteliali monostratificate cubiche alte (cellule follicolari). Sotto le cellule follicolari vi è la membrana pellucida (formata da mucopolisaccaridi) ed al centro vi è una grossa cellula uovo con un diametro di circa 150 mm (è una delle più grosse cellule dell'organismo). Nelle donne il numero di cellule uovo è già stabilito fin dalla alla nascita, varia da 500 mila a circa 1.000.000 per ovario e varia da donna a donna. L'uovo umano è oligolecitico , cioè è uno dei più piccoli uova che si conoscano nel regno animale e vegetale. Si deve però distinguere l'ovario prepuberale da quello puberale. • • L'ovario prepuberale ha solo follicoli primordiali. Il follicolo è formato da piccole cellule appiattite, con la membrana pellucida sottilissima. La cellula uovo ha un diametro di 120 mm. L'ovario puberale ha invece follicoli oofori primari che mediante la follicologenesi si trasformeranno. Prof. A. F. Cannone 6 Anatomia Umana OVOGENESI Il processo meiotico avviene in due fasi: fase cariocinetica (meiosi I) e fase riduzionale (meiosi II). Mentre nella fase cariocinetica si formano due cellule diploidi, nella fase riduzionale si formano 4 cellule apliodi. La cariocinesi, durante la vita embrionale, avviene solo negli ovogoni e non negli spermatogoni. L'ovogenesi degli ovogoni avviene all'interno degli ovari quando ancora il feto si trova nel grembo materno. Alla nascita, la bimba ha un certo numero di ovogoni (diploidi) che rimangono tali fino al momento della pubertà. Solo da questo momento, quando il follicolo inizia a maturare si trasformano in aploidi (avviene la riduzione, la II fase meiotica). Però non tutti gli ovuli vanno incontro a maturazione, la maggior parte va incontro ad una involuzione, diventano atresici e vengono eliminati. Delle circa 500 mila ovuli per ovario che si hanno alla nascita, solo il 10-15 % maturerà e di questi molti si atrofizzeranno. Infatti ad ogni ovulazione maturano una ventina di ovuli, solo uno maturando più degli altri produce degli ormoni che inibiscono gli altri ovuli che formeranno la corpora obligatia. TIPI DI OVOCITI L'uovo umano è oligolecitico (la lecitina è una sostanza fosfolipidica presente in tutti i tipi di uova, animali e vegetali), cioè è uno dei più piccoli uova che si conoscano nel regno animale e vegetale. Oligolecitiche (uovo piccolo, povero di lecitina) Mammiferi Mesolecitiche (uovo medio con poca lecitina) Pesci Megalecitiche o telolecitiche (uovo grosso, ricco di lecitina) Rettili, Uccelli Prof. A. F. Cannone 7 Anatomia Umana Ciclo ovarico Il ciclo ovario si svolge in 28 giorni. Tale periodo si può suddividere in due fasi: follicolare = maturazione del follicolo ooforo (follicologenesi); gestazionale = creazione delle condizioni affinchè il follicolo ooforo una volta maturato possa essere fecondato e quindi possa iniziare una gravidanza. Ogni fase si svolge in 14 giorni. Nella pubertà il follicolo primario in seguito alla produzione di FSH si evolve (follicologenesi) diventando progressivamente molto grosso (la cellula uovo rimane dalla stessa grandezza) e si trasforma in follicolo secondario. Il follicolo secondario è più grosso per due motivi: a) aumenta il numero delle cellule follicolari; b) le cellule follicolari producono una sostanza liquida (l'ormone 17-β estrandiolo) che rimane immagazzinata dentro gli spazi all'interno del follicolo, che dopo si trasforma in follicolo vescicoloso di Graaf. La fase finale della follicologenesi si ha il 14° giorno quando all'interno del follicolo secondario si forma una cavità grandissima che può arrivare anche a 2 cm. Questo è il follicolo vescicoloso di Graaf (di solito ad ogni ciclo si forma un solo follicolo di Graaf) nel cui interno vi è la cellula uovo, il liquor follicoli (è formato da un ormone, il 17-β estrandiolo) ed all'esterno vi è un rivestimento di cellule follicolari chiamato granulosa. La granulosa è rivestita da 2 teche, una interna che guarda verso la cavità del follicolo ed una esterna che guarda verso uno strato giallo dell'ovaio. Sulla teca esterna vi è la pregna vascolare, una struttura connettivale fortemente vascolarizzata Nella fase finale di maturazione la teca esterna si modifica in granulosa gialla (corpo luteo). Il colore giallo è dato dalla presenza del colesterolo. La follicologenesi termina il 14° giorno con l'ovulazione, infatti per la pressione interna esercitata dall'uovo, il follicolo di Graaf si avvicina alla superficie dell'ovario fino a rompere la parete dell'ovario e a scoppiare: a) il liquor follicoli (liquido follicolare) va nella cavità peritoneale e da qui passa nel circolo sanguigno; b) la cellula uovo con tutte le cellule follicolari del follicolo di Graaf cade nell'ampolla tubarica, cioè ovula (ovulazione). Nella tuba di Falloppio, può incontrare uno spermatozoo ed essere fecondata, oppure, se non viene fecondata, dopo 14 giorni viene espulsa con le mestruazioni. CICLO DEL COLESTEROLO La teca esterna dell'ovulo, durante l'ovulazione, si trasforma in corpo luteo di colore giallo. Il colore giallo è dato dalla presenza del colesterolo che forma gli ormoni sessuali (steroli): estrogeni, progesterone e testosterone. Gli steroli sono composti organici a struttura policiclica con all'interno il ciclopentanoperitrofenantrene a 27 atomi di carbonio al quale è unita la molecola dell'acido proprionico. La trasformazione del colesterolo in ormoni avviene allo stesso modo sia nell'ovario che nel testicolo. Il colesterolo in questi organi viene demolito fino a trasformarsi in ormoni sessuali. Nell'ovaio prima si produrrà testosterone e poi (per mezzo di enzimi) si produrranno estrogeni a 18 atomi di carbonio. Solo una piccola parte rimarrà come androgeni (3-4 %) il resto si trasformerà in estrogeni. È questo il motivo per cui nel corpo luteo vi sono anche androgeni. Nei testicoli invece il ciclo si arresta a 19 atomi di carbonio (androstano), quindi nell'uomo la maggior parte del prodotto è il testosterone. Solo una piccola parte di androstano, per azione dell'enzima aromatasi, si trasforma in estrano e da questo si formano gli estrogeni. Prof. A. F. Cannone 8 Anatomia Umana Ciclo uterino o endometriale I primi tre strati dell'endometrio, durante il ciclo uterino sono soggetti a modifiche: Fase rigenerativa dell'endometrio (primi giorni del ciclo) Fase proliferativa (Æ 14° o estrogenica giorno) dell'endometrio Strato germinativo basale con Membrana basale cellule stromali in ricrescita Epitelio monostratificato cubico Lo strato germinativo viene Membrana basale alto + ghiandole tubulari + progressivamente sostituito da addensata capillari spiralati Æ a. spirale Æ un sottile strato spugnoso a. iliaca interna Fase progestinica endometriale (14° Æ 28° giorno) Epitelio si trasforma in mucoso + Spesso strato spugnoso ghiandole tubulari (muco) + (formato dal progesterone) capillari spiralati Fase regressiva o desquamazione endometriale (verso 28° giorno) Epitelio mucoso + ghiandole tubulari (muco) + capillari spiralati Mestruazione (28° giorno) Membrana basale Viene eliminato lo strato Membrana basale spugnoso per la progressiva diminuzione di estrogeni e di progesterone Membrana basale Ciclo vaginale Le cellule dello strato epiteliale mucoso della vagina, durante i 28 giorni del ciclo ovarico, vengono modificati dalla presenza degli ormoni femminili: Fine mestruazione La dequamazione delle cellule endoteliali è scarsa con cellule esfoliate, precorneificate, poligonali. Pochi leucociti polimorfonucleati. Fase follicolare o estrogenica Poche cellule desquamate, piatte o ripiegate con molte le cellule corneificate con nuclei omogenei. Pochi leucociti polimorfonucleati. Fase luteica o progestinica Molte cellule desquamate, angolate o ripiegate con poche cellule corneificate con nuclei vescicolari. Molti leucociti polimorfonucleati. Fase premestruale Molte cellule desquamate. Si notano gruppi serrati di cellule precorneificate con nuclei vescicolari. È presente muco e molti leucociti polimorfonucleati. Le varie fasi del ciclo vaginale su esposte, si possono riconoscere osservando le cellule desquamate prelevate dalla vagina con uno striscio vaginale. • • Le cellule desquamate provengono dallo strato epiteliale mucoso I linfociti provengono dai sottostanti capillari vaginali. ORMONI CHE INTERAGISCONO NEI CICLI FEMMINILI Il 17 beta estrandiolo (estrogeno) si incomincia a formare il I° giorno del ciclo, per azione dell'FSH. Per mezzo dei capillari sanguigni passa nello stroma ovarico e da qui dentro il follicolo secondario che si trasforma in follicolo di Graaf. È per questo motivo che nel sangue delle donne la quantità di estrogeni è costante (poca) ed aumenta solo quando scoppia il follicolo di Graaf e si riversa in circolo. Prof. A. F. Cannone 9 Anatomia Umana La gonadotropinna follicolostimolante (FSH) ipofisaria attiva la crescita del follicolo primario in secondario. È solo dopo la stimolazione dell'FSH che le cellule follicolari incominciano a proliferare e a produrre estrogeni. (il target per l'FSH è la cellula follicolare). La quantità di FSH è massima il I° giorno del ciclo e dal 14° giorno in poi diminuisce progressivamente, per poi aumentare verso il 28° giorno. La progressiva diminuzione è un caso di feed-back negativo, perché la rottura del follicolo di Graaf e l'immissione in circolo di estrogeni, fa arrivare all'ipofisi il segale di inibire la produzione di FSH. Quando gli estrogeni (I° giorno del ciclo) sono pochi., il feed-back è positivo e si riforma FSH. La produzione dell'ormone luteinizzante LH è stimolata dalla presenza di estrogeni. L'LH stimola nell'ovaio la produzione di progesterone (14° giorno) e fa formare il corpo luteo. La produzione di LH non è costante, ma pulsante. La caduta avviene in maniera dolce. Il progesterone passa nel sangue e da qui arriva nel diencefalo fino ad inibire per feed-back negativo la produzione di LH ed invece eccitare per feed-back positivo la produzione di FSH. La quantità alta del progesterone si mantiene costante nel corpo luteo durante i 14 giorni finali del ciclo, ciò è importante perché assicura l'attecchimento del embrione. Il progesterone prodotto dal corpo luteo gravidico, induce la trasformazione dell'endometrio in decidua. La decidua a sua volta formerà la componente materna della placenta. Prof. A. F. Cannone 10 Anatomia Umana APPARATO GENITALE MASCHILE È formato da organi primari (testicoli) e organi accessori (dotti secretori, vescichette seminali, prostata, ghiandole bulbo-uretrali, pene) TESTICOLI I testicoli sono formati da due parti: il didimo e l'epididimo. Sono contenuti in uno scroto, esterno all'addome. Ogni testicolo è rivestito dalla tonaca albuginea. All'interno vi sono i tubuli seminiferi tra i quali si trovano le ghiandole interstiziali. I tubuli seminiferi sono separati fra di loro dai setti albuginei (connettivo capillare). I setti formano le logge (circa 250) ognuna delle quali è completamente riempita da tubuli seminiferi. Ogni loggia ha un sistema tubulare che va a finire nei tubuli retti che si anastomizzano e formano la rete testis e poi mediante i condotti efferenti vanno a finire nella testa dell'epididimo. All'epididimo segue il dotto deferente o funicolo spermatico, che mediante il canale inquinale entra nello scroto. Da qui passa nella cavità pelvica e termina nelle vescichette seminali. Nelle vescichette si uniscono i tubuli provenienti dai due testicoli e passano nella prostata e poi nell'uretra. L'uretra percorre il pene, un organo cilindrico contenente i corpi cavernosi, muscoli erettili, pieni di vasi venosi. Il pene serve sia per l'eliminazione dell'urina, sia per l'eiaculazione degli spermatozoi. DIDIMO Tubuli seminiferi Ghiandole interstiziali EPIDIDIMO Prof. A. F. Cannone Corpo, testa, coda Membrana basale (connettivo lamellare con fibre eleastiche) + cellule del Sertoli (testis barrier, ABP, attiva testosterone) + epitelio seminifero (spermatogoni, spermatociti, spermatidi, spermatozoi) Vasi sanguigni + vasi linfatici + cellule poligonali + cellule inerstiziali o intertubulari di Leyding (grosso nucleo sferico, granuli di Rencke, lipidi, testosterone attivo) Tubuli con cellule epiteliali ciliate (stereociglia) pseudostratificate 11 Anatomia Umana TUBULI SEMINIFERI I tubuli seminiferi sono delimitati esternamente da tessuto connettivale lamellare contenente fibre elastiche (membrana basale). Su questo strato si poggiano le cellule di sostegno o cellule del Sertoli e l'epitelio germinale contenente spermatogoni, spermatociti, spermatidi ed infine spermatozoi maturi e flagellati. CELLULE DEL SERTOLI Le cellule del Sertoli hanno un nucleo vescicoloso, il citoplasma contiene molti granuli e gocce lipidiche che aumentano con il progredire dell'età. Sono grosse cellule con la base ampia e l'apice rivolto verso il lume del tubulo. Alle cellule del Sertoli si attaccano gli spermatozoi, per prendere nutrimento. Il numero delle cellule del Sertoli rimane costante nell'adulto (non si ha mitosi). Sono molto resistenti alle radiazioni ionizzanti e a varie sostanze tossiche. Le cellule del Sertoli sono collegati fra di loro da sistemi di giunzione di tipo desmosomico (gap-junction). Tali giunzioni dividono la parete del tubulo in due compartimenti: basale ed adluminale Contiene gli spermatogoni e gli stati meiotici iniziali (leptotene) Compartimenti Basale della parete Adluminale Contiene gli stati avanzati della spermatogenesi (spermatociti e tubulare spermatidi) Sostegno meccanico ( testis barrier) alle cellule germinali. Hanno un ruolo nella liberazione degli spematozoi del tubulo Funzioni cellule del Sertoli seminifero. Regolano la spermatogenesi, determinando un andamento ciclico (inibina) e le fasi del processo differenziativo (FSH Æ testosterone). Scambio metabolico fra liquido tubulare e liquido interstiziale Funzione autoimmune (non fa arrivare il sangue negli spermatozoi) Fagocitano i corpi residuali dello spermatozoo Prof. A. F. Cannone 12 Anatomia Umana EPITELIO SEMINIFERO Osservando la distribuzione delle cellule germinali in un tubulo seminifero si vede che vi sono diverse associazioni cellulari che si succedono nel tempo una dopo l'altra : Periferia del tubulo sulla lamina basale Centro del tubulo Spermatogoni Spermatociti I e II, spermatidi, spermatozoi. La diversa stratificazione delle cellule germinali o ciclo dell'epitelio seminifero è dovuto a tre eventi: La spermatogenesi ha un carattere ciclico; La spermatogenesi avviene a velocità costante nel tempo; In ogni sezione trasversale si sviluppano contemporaneamente cellule germinali sfasati tra di loro di un intervallo di tempo costante. GHIANDOLE INTERSTIZIALI Le ghiandole interstiziali si trovano fra i vari tubuli seminiferi. Sono formate da vasi sanguigni e linfatici e da cellule connettivali interstiziali o intertubulari o di Leyding, contenenti grandi gruppi di cellule poligonali. Le cellule interstiziali di Leyding hanno in grosso nucleo sferico a volte in posizione eccentrica. Il citoplasma contiene molti granuli (cristalli di Renche) alcuni di natura lipidica. Queste cellule vanno incontro a processi di degenerazione indicativi di una funzione secretoria. Producono infatti testosterone e recettori di superficie. Eventuali eccessi di testosterone presente nei tubuli seminiferi, vengono incamerati dalle cellule del Sertoli, le quali per mezzo dell'enzima acromatasi trasformano il testosterone in eccesso in estrogeni non attivabili (cioè che non si possono utilizzare). Tali estrogeni non attivabili si accumulano nel reticolo endoplasmatico (infatti il reticolo è molto sviluppato). EPIDIDIMO L'epididimo è formato da un corpo, una testa e una coda. All'interno vi sono una serie di tubuli formati da cellule cilindriche alte (hanno il nucleo in posizione alta) o basse (hanno il nucleo in posizione bassa). L'insieme da la sensazione che vi sono più strati cellulari, invece vi è un solo strato di tessuto epiteliale pseudostratificato che poggia sulla membrana basale. Le cellule sono ciliate (stereociglia o cellule a pennacchio). Prof. A. F. Cannone 13 Anatomia Umana SPERMATOZOI Lo spermatozoo maturo è formato da un corpo cellulare e da un acrosoma (che con i suoi enzimi serve al riconoscimento specie-specifica e anche per poter bucare l'uovo durante la fecondazione) e da un flagello (che serve per poter penetrare nelle vie genitali femminili). Lo spermatozoo però quando si forma è ancora incapace a fecondare, ha bisogno di un processo di capacitazione, dopo può percorre le vie spermatiche e le vie genitali femminili e poi può fecondare. La formazione dell'acrosoma avviene in diversi fasi della spermiogenesi: L'apparto del Golgi dello spermatogonio inizia ad ipertrofizzarsi e a livello dei dittiosomi si incominciano a sintetizzare tutte le proteine che serviranno al riconoscimento specie-specifico dell'uovo (iarulonidasi) e alla lisi (enzimi litici) degli apparati che vi sono attorno all'uovo (le cellule follicolari e la sostanza pellucida)., quando avviene la fecondazione. Fase del Golgi Fase cappuccio acrosomico del Si forma come una vescicola che ingrandendosi riveste i due terzi anteriori del nucleo, tale vescicola è il cappuccio acrosomico. I due centrioli migrano ai poli opposti: uno si mette vicino al nucleo e uno funziona da corpuscolo basale per il flagello che si sta formando. Fase acrosomiale Fase maturazione Il nucleo si allunga e l'apparato acrosomiale si adatta alla forma del nucleo. I mitocondri e il citoplasma si dispongono sotto il nucleo e formano lo spilema o collo e sotto si forma il flagello. di Lo spermatozoide completa la sua trasformazione in spermatozoo, il nucleo e l'acrosoma assumono la forma caratteristica della specie. Il flagello al microscopio elettronico si vede formato da 9 coppie di microtubuli radiali ed una coppia centrale. Tali microtubuli servono alla contrazione. I microtubuli sono formati da tubulina (proteina costituita da actomiosina). Nello spilema e nel segmento centrale, attorno ai microtubuli vi sono delle fibre esterne. Sullo spilema vi è una guaina ricca di citoplasma e di mitocondri. Prof. A. F. Cannone 14 Anatomia Umana CAPACITAZIONE SPERMATICA Per capacitazione si intendono i processi che rendono lo spermatozoo capace a fecondare. La capacitazione dello spermatozoo inizia con la spermiogenesi. In tale fase, lo spermatozoo assume (l'apparto del Golgi, il cappuccio, l'acrosoma) la forma tipica e le caratteristiche della specie. Nell'epididimo lo spermatozoo diventa maturo e mobile, perché in questa zona vi è ATP. Dopo, lo spermatozoo sosta nelle vescicole seminali. Qui vi è un liquido ricco di fruttosio ed un pH ottimale che conferiscono energia e vitalità allo spermatozoo. Prima di essere eiaculato lo sperma si arricchisce del secreto di altre due ghiandole annesse all'apparato urogenitale: la prostata e le ghiandole bulbo-uretrali. Queste ghiandole hanno il compito di formare un liquido lubrificante, utile nella copula. Lo sperma va a finire nella vagina ed anche qui proseguono i processi di capacitazione. La maggior parte degli spermatozoi va incontro a fenomeni degenerativi a causa del pH vaginale acido (per la presenza di acido lattico prodotto dai batteri acidofili vaginali). Altri spermatozoi vengono fagocitati dai macrofagi. Altri, gli ultimi spermatozoi ad essere eiaculati, essendo molto alcalini vengono risucchiati nell'utero. Qui, la maggior parte degli spermatozoi rimasti, viene fagocitata dalle cellule dell'endometrio uterino. Nella tuba, lo spermatozoo completa la capacitazione in quando è capace di riconoscere l'ovulo della stessa specie. Spermatogenesi Acquista la forma e le caratteristiche della specie Epididimo (ATP) Diventa maturo e mobile Vescicole seminali (fruttosio + pH Acquista energia e vitalità ottimale) Prostata e ghiandole bulbo-uretrali Acquista un liquido lubrificante utile alla copula Vagina Alcuni spermatozoi degenerano, altri vengono fagocitati, solo alcuni (alcalini) vengono risucchiati nell'utero Utero Vengono fagocitati dalle cellule dell'endometrio (macrofagi vaginali + granulociti neutrofili) Tuba Riconoscimento specie-specifica PERCORSO SPERMATICO Alla fine della spermiogenesi lo spermio è quasi privo di citoplasma ma ha dei corpi residuali che vengono fagocitate dalle cellule del Sertoli. Gli spermi si liberano dalle cellule del Sertoli ed escono nel lume del tubulo seminifero, poi passano nei tubuli retti e da qui nella rete testis (testa dell'epididimo) e attraverso l'epididimo raggiungono i condotti deferenti. All'interno di questi condotti, essendosi maturati, passano mediante il canale inquinale nello scroto e da qui nelle vescichette seminali, nella prostata, nell'uretra ed attraverso il pene vengono eiaculati. Lungo il percorso degli spermatozoi si aggiungono i secreti prodotti dalle vescichette seminali, dalla prostata, dalle ghiandole bulbo -uretrale. Tali secreti formano il liquido seminale che unendosi agli spermatozoi forma lo sperma. Il moto degli spermi nelle vie genitali avviene sia perché lo spermio ha una coda mobile, sia per i moti peristaltici dei muscoli dei dotti escretori. Lo spermatozoo in seguito alla eiaculazione, va a finire all'interno della vagina, ed in seguito a contrazioni ritmiche dell'utero e delle trombe uterine viene risucchiato nella cavità uterina e successivamente nelle trombe uterine. Circa un'ora dopo la penetrazione in vagina, gli spermatozoi arrivano nell'ampolla tubarica. Gli spermatozoi eiaculati nelle vie genitali femminili sono circa 200-300 milioni. Dalla vagina, attraverso il canale cervicale passano nell'utero e poi attraverso le giunzione utero-tubarica, nella tuba. Però la maggior parte degli spermi viene fagocitata dai granulociti neutrofili e dai macrofagi che si trovano nella vagina e nell'utero. Solo pochi spermi arrivano nella tuba entro pochi minuti della deposizione del seme nella vagina, altri continuano il flusso lungo la tuba per alcuni ore e attraverso l'ampolla si riversano nella cavità peritoneale. Se lo spermio non incontra l'ovulo, muore dopo circa 24 ore. Se invece incontra l'ovulo, uno solo entra ed inizia la fecondazione, i pochi spermi rimasti all'esterno vengono attaccati dalle spermolisine, sostanze prodotte dalle cellule follicolari dell'ovulo, ed uccisi. Con la fecondazione iniziano tutti quegli eventi moleclari che portano alla segmentazione e quindi all'embriogenesi. Prof. A. F. Cannone 15 Anatomia Umana Tragitto dello spermatozoo nelle vie spermatiche e nelle vie genitali femminili Tessuto germinale Æ tubuli seminiferiÆ tubuli rettiÆ rete testisÆ epididimoÆ dotto deferenteÆ canale inquinale (scroto ) Æ (cavità pelvica) vescichetta seminaleÆ ( si uniscono i dotti di entrambi i testicoli) Æ prostataÆ ghiandole bulbo-uretraleÆ uretraÆ pene Æ ……….. Æ vagina Æ utero Æ ampolla tubarica Æ cavità peritoneale (legamento largo dell'utero) SPERMIOGENESI La spermiogenesi e quel processo di modificazioni strutturali e metaboliche cui va incontro lo spermatogonio nella trasformazione a spermatozoo. La spermatogenesi viene attivata dall'ormone follicolo stimolatore (FSH) proveniente dall'ipofisi. L'FSH agisce sulle cellule del Sertoli (targhet). Vediamo come arriva nelle cellule del Sertoli. L'FSH nel sangue trova una proteina, l'SHBG (Sieroide-Hormon-binding-Globulin) e si fa trasportare negli spazi interstiziali del testicolo, nelle cellule di Leyding. Le cellule del Leyding, stimolate, iniziano a produrre testosterone inattivo, il quale mediante una proteina (recettore specifico di superficie, prodotta dalle cellule del Sertoli, l'ABP Antrogen-BindingProtein), si attiva. Il testosterone attivato, entra nello spermatogonio (epitelio germinale). Dentro il citoplasma dello spermatogonio trova un recettore, la 5-α-reduttasi che trasforma il testosterone in 5-α-deidrotestosterone forma attiva del testosterone). Il 5-α-deidrotestosterone entra nel nucleo dello spermatogonio, e fa avvenire la fosforilazione delle proteine istoniche e quindi lo scioglimento delle 2 eliche del DNA e l'inizio di tutte quelle attività che fanno trasformare lo spermatogonio in spermatozoo. Dopo la formazione dello spermatozoo, le cellule del Sertoli producono una certa quantità di un peptide, l'inibina che blocca la spermatogenesi di altri spermatogoni. Quando l'inibina si esaurisce (l'inibina esercita un feed back nei confronti dell'FSH) l'FSH inizia un nuovo ciclo.La spermatogenesi si suddivide in tre fasi: mitotica (maturazione degli spermatogoni),meiosi (spermatociti I diploidi, spermatociti II aploidi), spermiogenesi (spermatidi, spermatozoi). Gli spermatogoni (2n) quando vengono attivati dalla 5-α-reduttasi iniziano la spermatogenesi che in genere dura 8-10 giorni. Gli spermatogoni prima di entrare in meiosi si suddividono per mitosi e formano spermatogoni di riserva (nucleo pallido) per successive spermatogenesi e altri spermatogoni (nucleo scuro) che si evolvono rapidamente per divenire spermi. Questi spermatogoni per mitosi si suddividono in spermatociti I (fase di accrescimento) in cui il nucleo (2n) si presenta allo stadio di leptotene, cioè i cromosomi sono dipanati prima di iniziare la meiosi I. Gli spermatociti I (hanno cromosomi omologhi in cui sono appaiati quello materno e quello paterno) sono legati insieme da un ponte citoplasmatico perché la citodieresi non si è ancora attivata. Quando inizia la meiosi I gli spermatociti I si portano dalla membrana basale verso gli strati più interni dell'epitelio seminifero. Incomincia anche la differenziazione dell'acrosoma. Gli spermatociti I si trasformano in spermatociti II (siccome gli spermatociti I non hanno duplicato il corredo cromosomico i due spermatociti II sono di conseguenza aploidi). In questa fase la differenziazione dell'acrosoma si fa più evidente (il citoplasma si riduce molto). Gli spermatociti II (n) per meiosi II si dividono e si formano gli spermatidi (il citoplasma è quasi assente) da questi attraverso un processo di trasformazione (spermiogenesi) si formano gli spermatozoi nel quale si differenziano strutture come il flagello e l'acrosoma che intervengono nella fecondazione. Prof. A. F. Cannone 16 Anatomia Umana FECONDAZIONE Lo spermio per fecondare l'ovulo, prima deve perforare le cellule follicolari, poi la membrana pellucida, dopo la membrana cellulare ed infine può penetrare nell'ovulo. Sulla testa dello spermatozoo vi è l'acrosoma che oltre a contenere enzimi litici, contiene proteine in grado di conoscere la specificità dell'uovo. Infatti sulla superficie dell'ovulo vi sono dei gruppi o siti aptenici (di attacco) che permettono questo riconoscimento specie-specifica fra i due gameti. Gli apteni si trovano alla estremità dei prolungamenti che partono dalla cellule uovo e oltrepassano la zona pellucida. Quando uno spermatozoo si lega con il suo recettore specifico ad un aptene, gli enzimi litici dell'acrosoma dello spermio, aprono un varco (reazione acrosomiale) nelle cellule follicolari ed avviene una reazione corticale sulla superficie dell'ovulo, che ritira immediatamente tutti i prolungamenti (apteni) ed anche lo spermatozoo che vi è legato, e non fa entrare altri spermatozoi. A volte possono essere ritirati due o più spermatozoi, ma quello che feconda l'ovulo è solo uno, gli altri vengono distrutti dai lisosomi ovulari. La reazione corticale consiste nella inattivazione dei recettori dello spermatozoo e nel suo indurimento (vengono agglutinati gli enzimi proteolitici), per mezzo di enzimi secreti dalla membrana pellucida. Dopo la reazione corticale, l'ovocita, all'ingresso dello spermatozoo completa la II divisione meiotica e forma una cellula uovo normale e un polocita. Cioè la cellula uovo si prepara ad essere fecondata. EVENTI MOLECOLARI DELLA SEGMENTAZIONE I ° SETTIMANA, DECIDUAZIONE DELL'ENTOMETRIO Appena l'uovo è fecondato, forma l'HMG che ha azione immunosoppressiva, per evitare che lo zigote venga espulso. I primi due blastomeri vengono attivati meglio modulati (indirizzati STADIO ad una certa segmentazione, ad un certo volume…) da sistemi di controllo che nel nostro caso DIDERMICO sono dei neurotrasmettitori (sono delle sostanze per cui il S.N.C. e il S.N. neurovegetativo riesce a controllare tutte le attività del nostro corpo. I neurotrasmettitori che intervengono sono tutti monoaminergici: acetilcolina, catecolammina, (adrenalina, noradrenalina, FILOGENESI ED ONTOGENESI dopamina), serotonina. I primi neurotrasmettitori che agiscono sono l'acetilcolina e la serotonina. Dopo agiscono le catecolammine. Più si sviluppa l'embrione più neurotrasmettitori agiscono. CIRCOLAZIONE MATERNO FETALE PRIMA SETTIMANA I settimana (segmentazione, trasporto, impianto, decidualizzazione dell'endometrio). Caratteristiche utrastrutturali della I fase di sviluppo: Le uova che contemporaneamente maturano sono circa 20 di questi solo uno è più grosso degli altri perché produce gli estrogeni. Gli altri si atrofizzano. A volte quelli che si ingrossano e maturano sono più di uno. Nascono in questo caso, se fecondati, i gemelli non monoovulari (bi-triovulari….) invece la fecondazione bispermica, trispermica porta sempre alla nascita di un bambino monoovulare. Se invece un normale uovo fecondato si divide in due blastomeri e questi invece di proseguire la segmentazione insieme, la proseguono ognuno per conto proprio, in questo caso si formeranno dei gemelli monoovulari o identici ( cioè hanno lo stesso corredo cromosomico) perché si sono sviluppati dallo stesso ovulo. Gli spermatozoi entrano con tutta la coda nel citoplasma dell'uovo (la coda non esercita nessuna funzione nella riproduzione e nello sviluppo ma degenera e scompare rapidamente). L'uovo, dopo essere stato fecondato nella ampolla uterina (fusione dei gameti e delle membrane plasmatiche), avvolto dalla membrana pellucida, si chiama zigote. Con la fecondazione inizia la segmentazione. La cellula uovo misura circa 250µm di diametro, non è gigantesca. I primi 2 blastomeri che si formano saranno di 175 µm. I secondi 4 blastomerisaranno di 8 µm e così via fino a che le cellule raggiungono la grandezza delle cellule embrionali della specie (stadio di 16 blastomeri). Il prodotto finale è la morula in cui vi sono cellule piccole che hanno poco citoplasma e scarse risorse energetiche. Nel frattempo l'endometrio diventa spugnoso perché si è iniziato a produrre progesterone e la mucosa endometriale raggiunge il massimo sviluppo (siamo al 5 - 6 giorno). Questa è una fase molto importante sia per l'embrione che per la gestante. Infatti spesso le aderenze dell'appendice ostacolano (stenosi) le trombe uterine che possono essere causa di sterilità; oppure lasciano passare lo Prof. A. F. Cannone 17 Anatomia Umana spermio ma non l'ovulo fecondato. In tal modo si può avere una gravidanza tubarica extrauterina molto pericolosa perché si verifica la rottura della tromba, emorragie, peritoniti e spesso anche la morte. La morula dopo il 5° giorno si vitalizza e diventa cava al centro, le cellule, prima indifferenziati, si differenziano, si accrescono e si dispongono in superficie attorno alla vescicola blastocistica o blastocisti. Tale fase della segmentazione si chiama blastula. La blastula scende nell'utero. Le cellule della blastula che si dispongono in superficie, sono formate da un solo strato di cellule piatte, lo trofoblasto (dette così perché attuano il trofismo, cioè la nutrizione dell'embrione), le cellule interne si accumulano in un polo e formano l'embrioblato (detto così perché daranno origine all'embrione ed anche all'aminios e al sacco vitellino). La blastula, verso il 7° giorno dall'ovulazione, arriva a contatto con l'endometrio uterino (l'endometrio è allo stato premestruale ed è quindi spugnoso a causa dell'azione degli estrogeni e del progesterone). A contatto con l'endometrio, i microsomi dello trofoblasto iniziano a formare la fosfatasi acida che prima distrugge le proteine della mucosa e poi fa aderire lo trofoblasto all'endometrio. Inizia così l'impianto di tipo interstiziale della blastula che per mezzo dello trofoblasto penetra nello stroma sottostante riccamente vascolarizzato. Lo trofoblasto rompe i vasi (i capillari uterini derivano dall'arteria uterina, la quale deriva da un ramo interno dell'aorta addominale) e vi prolifera dentro fino ai vasi sanguigni, formando prima il chorion e dopo le porzioni fetali della placenta. La blastocisti continua ancora a penetrare nell'endometrio, mentre lo trofoblasto continua a proliferare. Si formano 2 strati: uno strato monostratificato (monocellulare o cellule d'impianto) interno o citotrofoblasto ed uno sinciziale (marginale, formato dalle cellule di Langhans) esterno o sinciziotrofoblasto. Il sinciziotrofoblasto non è un sincizio (cellula polinucleata che si forma da singole cellule che fondono la loro membrana e si mettono insieme) ma è un plasmodio (cellula polinucleata che si forma in quando alla divisione del nucleo non segue la divisione del citoplasma e la formazione di due cellule). Durante le prime tre settimane (21 giorni) il sinciziotrofoblasto proliferando (verso la decidua) forma i villi secondari che a seconda della forma si dividono in: chorium frondosum (sono villi coriali molto ramificati) e chorium laeve o liscio (sono villi coriali poco sviluppati e lisci. Si formano dalle protuberanze che si allontanano dal frondosum, quando tali protuberanze diventano sottili e lisci). I villi secondari sono formati da tre strati cellulari: internamente dal mesenchima extraembrionale, sopra dal citotrofoblasto ed esternamente dal sinciziotrofoblasto. Il citotrofoblasto invece emette delle propaggini (villi primari ad un solo strato cellulare che sono dirette alle lacune ematiche). Le cellule del citotrofoblasto diminuiscono progressivamente di numero con l'avanzare della gravidanza e difficilmente si trovano dopo il terzo mese. I villi primari e secondari non contengono vasi sanguigni, ma traggono nutrimento per osmosi dal sangue materno. Il chorion è formato dall'insieme dei villi coriali e dallo stroma connetivale. La blastocisti ad un certo punto si invagina tutta nell'endometrio ed avviene la decidualizzazione dell'endometrio. Sopra la blastocisti si riforma l'endometrio ed il muco, nel frattempo le proliferazioni trofoblastiche si trasformano in villi coriali, penetrano nello stroma (connettivo), raggiungono l'endotelio e pescano nel sangue uterino. I fibroblasti del connettivo reagiscono all'azione litica dei villi e si trasformano in grosse cellule rotondeggianti e polipoidi dette cellule deciduali. Queste cellule circondano i villi e le lacune ematiche dell'endometrio e modificano la mucosa uterina in modo spugnoso, la mucosa uterina così modificata si chiama decidua (è il progesterone ad indurre la trasformazione dell'endometrio in decidua. Il progesterone durante la gravidanza viene prodotto dalla placenta e non dal corpo luteo. Il progesterone serve anche al buon proseguimento della gravidanza). La reazione deciduale serve a migliorare la nutrizione dell'embrione e serve anche a proteggere la mucosa uterina dall'ulteriore azione litica dei villi. La reazione deciduale inizia nella regione uterina a contatto con il corion frondosum, in cui si forma la decidua basale. Poi si estende con il corion liscio che forma la decidua capsulare. Più tardi interesserà la rimanente parte dell'endometrio e forma la decidua parietale (marginale), con il proseguire della gestazione la decidua capsulare e la parietale si fonderanno, invece la decidua basale saldandosi al corion frondosum formerà la placenta (il corion costituirà la componente fetale della placenta, invece la decidua basale, la componente materna). Prof. A. F. Cannone 18 Anatomia Umana STADIO DIDERMICO La blastocisti, quando entra nell'endometrio, esternamente è formata dallo trofoblasto (cito e sinciziotrofoblasto), da un ebrioblasto e da una cavità blastocistica detta celoma extraembrionale. L'embrioblasto a sua volta si suddivide in due strati cellulari o foglietti embrionali (stadio didermico), esternamente si forma l'ectoderma primitivo ed internamente l'endoderma primitivo (formato da cellule basse che formano il tetto del sacco vitellino). Entrambi questi foglietti parteciperanno alla formazione dell'embrione (disco embrionale). Le cellule ectodermiche a contatto con lo trofoblasto (il quale è a contatto con l'amnios) si chiamano amnioectodermiche. Lo strato ectodermico si scava all'interno e forma l'amnios (cavità amniotica) che circonda il sacco vitellino primario, formatosi dall'endoderma. Il sacco vitellino primario corrisponde all'antico blastocele, ed è circondato prima dal citotrofoblasto e con la formazione del mesenchima extraembrionale della splancnopleura. Il sacco vitellino definito si forma quando si dilata il celoma extraembrionale. Il sacco vitellino primario a sua volta forma un peduncolo mesenchimatico che all'estremità porta la cisti celomatica. FILOGENESI ED ONTOGENESI Nel corso dell'evoluzione, in molti tre miliardi di anni, i virus si sono trasformati in batteri, poi in vegetali ed animali, e si sono formati molti phila di esseri viventi. (filogenesi). Lo stesso accade all'embrione, quando si formano gli organi). Noi veniamo concepiti da cellule che non sono nemmeno cellule (gli spermi e gli ovuli sono aploidi). Infatti hanno un corredo cromosomico dimezzato. Da soli non possono sopravvivere, per cui hanno bisogno di legarsi insieme mediante la fecondazione. Quindi da cellule imperfette passiamo allo stadio di cellule eucariote (zigoti), il quale si segmenta e si trasforma in morula con cellule indifferenziate. Dopo inizia la fase della differenziazione e si producono miliardi di miliardi di cellule. Durante la nostra embriogenesi si passa da animali unicellulari (zigote) a pluricellulari (morula), ai molluschi (senza colonna vertebrale), ai cordati (animali con corda dorsale) e dopo si sviluppa lo scheletro che prima non è osseo ma cartilagineo come nei Selaci. Poi lo scheletro diventa osseo (vertebrati veri e propri). Quindi in 9 mesi si riesce a ripercorrere tutte le tappe dell'evoluzione degli esseri viventi. Secondo Heeckel l'ontogenesi ricapitola la filogenesi. La filogenesi (evoluzione degli esseri viventi, phila) e l'ontogenesi (evoluzione dell'embrione) sono due linee non parallele perché la filogenesi è una linea infinita, mentre l'ontogenesi è finita. Quindi quando si parla di ontogenesi si riferisce solo a una parte evolutiva della filogenesi. L'ontogenesi inizia dal gamete che si incontra con l'altro eterogamete e forma lo zigote. Fino alla morula, siamo uguali a tutti gli altri animali. Dalla blastula in poi inizia la vera evoluzione dell'uomo in cui si differiscono i trofoblasti e l'embrioblasto. Dalle cellule dell'embrioblasto poi inizia la vera differenziazione. Infatti ogni cellula darà origine a un determinato organo, un determinato tessuto. CIRCOLAZIONE MATERNO FETALE Verso la fine della III settimana alcune cellule del mesoderma extraembrionale, dentro la placenta, si differenziano e formano le lacune (isole) ematiche in cui il sangue non coagula perché il trofoblasto ha anche un'attività antitromboplastinica, cioè inattiva la tromboplastina, che le permette di non far coagulare il sangue. Infatti il sangue fluido può scambiare meglio le sostanze nutritive con le cellule. Con la formazione delle lacune sanguigne inizia la fase della nutrizione diffusiva operata dal sangue materno sui tessuti embrionali. Quindi da una nutrizione autotrofa in cui la cellula utilizza il vitello, si passa alla nutrizione degli umori circolanti o istiotrofa, nella tuba (si utilizzano per osmosi i liquidi materni, tubarici) ed infine ad una nutrizione diffusiva o ematotrofa. Con la formazione del cordone ombelicale si ha poi una nutrizione vascolare.Verso la III settimana, il corion si vascolarizza (si forma la placenta) si sviluppa in cuore e un sistema circolatorio. I villi si ancorano saldamente alla decidua basale. Alla fine della IV settimana compaiono le connessioni fra vasi del corion (feto) e quelli della madre. Tale connessione è data dal peduncolo ombelicale che stabilisce una circolazione feto - placentale. All'interno del peduncolo embrionale vi sono i vasi collettori. I villi assorbono nutrimento per osmosi dal sangue materno e da qui passano nella vena ombelicale e dai vasi collettori, nell'embrione. I prodotti di rifiuto sono portati dal feto al sangue materno attraverso le due arterie ombelicali che sono i nomi delle due arterie ipogastriche fetali. Dalle arterie ombelicali, dai villi coriali le sostanze di rifiuto e CO2 passano nella rete ematica materna sempre per osmosi. I villi coriali sono ossigenati direttamente dal sangue materno. Nella placenta c'è una circolazione invertita come avviene nella piccola circolazione polmonare. Quindi la placenta si comporta come il polmone del feto, entro la vena ombelicale ricca di O2 ed escono 2 arterie ombelicali ricche di CO2. Dentro le lacune ematiche (il sangue è della placenta e quindi è sangue materno) vi arrivano le arterie materne con all'estremità le cellule di Offbawer che chiudono, stenosizzano (induriscono) le arterie. Quando le cellule di Offbawer si rilasciano, il sangue zampilla verso l'alto e si accumula in alto. In basso si accumula invece il sangue venoso che filtra dai sottili villi coriali (dentro hanno una arteriola e una venula fetale). Quindi pur non essendoci nessuna compartecipazione il sangue si divide ed essendo il sangue arterioso in alto i villi pescano dal sangue arterioso. Dentro le lacune vi è l'80% di sangue arterioso e il 20% di venoso. Quest'ultimo sangue poi fluisce dalle vene ed entra in circolo materno. Prof. A. F. Cannone 19 Anatomia Umana SISTEMA NERVOSO Il sistema nervoso è suddiviso in tre tipi: 1) il sistema nervoso centrale, che comprende l'encefalo ed il midollo spinale; 2) il sistema nervoso periferico, che comprende tutti i nervi che collegano il sistema nervoso centrale agli organi periferici. 3) il sistema nervoso vegetativo o autonomo o viscerale, che si origina dal sistema nervoso periferico, ma è involontario. MENINGI Il sistema nervoso centrale (encefalo e midollo spinale è ricoperto da tre meningi: 1) dura madre (a contatto con le ossa cranici e dello speco vertebrale); 2) aracnoide (membrana intermedia riccamente vascolarizzata); 3) pia madre (a contatto con l'encefalo ed il midollo spinale). Gli spazi fra le meningi e l'interno dei ventricoli cerebrali e l'ependima, vi è il liquido cefalo rachidiano che viene formato dall'aracnoide. Il compito del liquido cefalo rachidiano è: attutire i colpi al s.n.c., nutrire le cellule nervose e non, eliminare le cellule morte. Prof. A. F. Cannone 20 Anatomia Umana ENCEFALO L'encefalo si può suddividere in: - telencefalo = è la parte esterna, la sostanza grigia o corteccia, suddivisa in due emisferi (sotto vi è il corpo calloso) in cui si notano le scissure, i lobi e le circonvoluzioni. All'interno vi è il I e II ventricolo. Moltissimi sono le funzioni del telencefalo. - diencefalo = superiormente vi è il talamo ed inferiormente l'ipotalamo. All'interno si trovano l'ipofisi, l'epifisi ed il III ventricolo. - mesencefalo = si trova sotto il corpo calloso del telencefalo. Contiene i peduncoli che collegano il diencefalo al romboencefalo. Vi è pure l'acquedotto di Silvio che collega il III ventricolo al IV. - romboencefalo = contiene il ponte di Varolio (collega il cervelletto ed il bulbo al mesencefalo) ed il cervelletto. - mielencefalo o bulbo = si continua nel midollo spinale. Contiene i nuclei della respirazione, del battito cardiaco, della peristalsi intestinale. Dall'encefalo fuoriescono i nervi encefalici. Prof. A. F. Cannone 21 Anatomia Umana Funzioni dell’encefalo NERVI ENCEFALICI I nervi encefalici o cranici sono nervi o sensoriali (portano all'encefalo gli impulsi che provengono dai recettori) o motori (portano gli stimoli ai muscoli) o misti. I nervi cranici sono 12 paia : oculomotore, acustico, olfattivo, trigemino, acustico, abducente, glossofaringeo, accessorio, ipoglosso, trocleare, facciale, vago. Prof. A. F. Cannone 22 Anatomia Umana MIDOLLO SPINALE Si origina nel bulbo e decorre dentro al canale midollare. Al centro del midollo vi è il canale ependimale, pieno di liquido cefalo rachidiano che arriva anche nei ventricoli cerebrali. Esternamente vi è la sostanza bianca ed all'interno sostanza grigia disposta a forma di H con due corna anteriori e due posteriori. Dalle corna anteriori escono le fibre motrici che raggiungono, con i nervi motori, i muscoli. Alle corna posteriori giungono le fibre sensitive provenienti dai gangli spinali (dove vi sono i neuroni a T o bipolari che servono a far entrare e non ad uscire gli stimoli nervosi). Le fibre sensitive e motori si uniscono fuori dal midollo spinale e formano 31 paia di nervi spinali misti. NERVI SPINALI I nervi spinali sono 31 paia. suddivisi in 8 cervicali, 12 toracici, 5 lombari, 5 sacrali ed 1 coccigeo SISTEMA NERVOSO AUTONOMO O VEGETATIVO Il sistema nervoso autonomo si origina dai gangli simpatici che si formano dal corno anteriore del midollo spinale (motore). Tali gangli sono formati da neuroni pregangliari e postgangliari. Si suddivide in simpatico e parasimpatico. a) simpatico = è costituito da nervi che emergono dai gangli cervicali, toracici e lombari del midollo spinale. Tale sistema funziona mediante i mediatori chimici acetilcolina (sui neuroni pregangliari) ed adrenalina (sui neuroni postgangliari). La stimolazione del simpatico causa la costrizione di alcune arterie, accelerazione del battito cardiaco, aumento della glicemia, inibisce le contrazioni e secrezioni gastrointestinali, contrae lo sfintere analedilatazione delle pupille, dilata i bronchi, stimola la secrezione di adrenalina e di noradrenalina, rilascia la vescica e contrae lo sfintere uretrale. b) parasimpatico = è costituito dai nervi che emergono dai gangli bulbari e sacrali. Tale sistema funziona mediante l'acetilcolina che agisce sia sui gangli pregangliari che postgangliari.La stimolazione del parasimaptico causa la contrazione della pupilla, la stimolazione della salivazione, contrae i bronchi, rallenta il battito cardiaco, stimola l'attività gastrica, stimola la cistifellea, stimola la peristalsi e rilascia lo sfintere anale, contrae la vescica e rilascia lo sfintere uretrale. Prof. A. F. Cannone 23
Scaricare