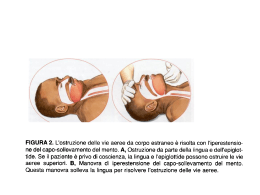
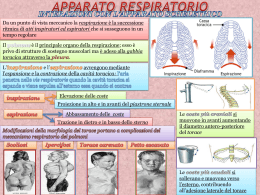
La regolazione della ventilazione • Compito del sistema respiratorio è quello di fornire ossigeno e rimuovere anidride carbonica dalle cellule ad una velocità adeguata a rispondere alle richieste metaboliche. • Quando ciò accade, nel sangue arterioso sistemico la PO2 e la PCO2 avranno valori pari a 100 mmHg e 40 mmHg, rispettivamente • Per mantenere questi valori, i meccanismi di controllo della ventilazione devono consentire un ritmo automatico che coordini la contrazione dei diversi muscoli respiratori al fine di regolare opportunamente la frequenza e il volume dell’atto respiratorio (da cui la ventilazione dipende) • In condizioni di eupnea si effettuano circa 13-15 atti respiratori al minuto • Tali atti continuano anche durante il sonno, quindi sono automatici • È possibile controllare volontariamente frequenza e durata dell’atto respiratorio o arrestare il respiro (apnea volontaria), ma tutte queste modificazioni volontarie hanno durata effimera: dopo poco tempo il respiro deve riprendere il suo andamento abituale • Evidentemente esistono dei meccanismi di controllo atti a riportare la funzione nell’ambito della norma • Il controllo della ventilazione avviene mediante due meccanismi: • Chimico • Nervoso • Entrambi concorrono al costante mantenimento della composizione dei gas respiratori nel sangue arterioso sistemico in tutte le situazioni fisiologiche: dall’esercizio fisico, al sonno, alla permanenza ad alta quota… Regolazione chimica del respiro • Durante un attacco d’asma, o in casi che provochino simili difficoltà respiratorie, si ha la sensazione di mancanza d’aria, nota come fame d’aria. • Sensazione simile si può provare rallentando volontariamente il respiro e cercando di mantenere costante la profondità di ciascun atto respiratorio • Dopo i primi secondi non si è più in grado di mantenere un ritmo respiratorio stabile perché esso diventa inevitabilmente più veloce e profondo • Ciò accade perché, alterando il pattern respiratorio, si altera la composizione chimica del sangue arterioso sistemico Regolazione chimica del respiro • I cambiamenti delle concentrazioni chimiche dei gas nel sangue vengono individuati da chemocettori localizzati nelle maggiori arterie e nel cervello • I chemocettori inviano poi segnali ai centri respiratori tramite neuroni afferenti • I centri respiratori rispondono variando la frequenza e la profondità del respiro per riportare le pressioni parziali dei gas nel sangue arterioso ai loro valori normali • Contemporaneamente i chemocettori inviano informazioni alla corteccia cerebrale evocando la sensazione cosciente di fame d’aria Variazioni delle concentrazioni chimiche E’ sufficiente un aumento del 4% del valore della PCO2 nel sangue arterioso per determinare un aumento della ventilazione da 5-7 l/min fino a 12 l/min (quasi il 100%) per favorire l’eliminazione dell’eccesso di CO2 Anche una diminuzione della PO2 porta ad un aumento della ventilazione mirato ad assumere più O2 dall’ambiente, ma per aversi un significativo aumento della ventilazione polmonare, la PO2 deve scendere sotto il 12%. Quindi la ventilazione è più sensibile alle variazioni della PCO2 che non della PO2 Infine anche alterazioni del pH ematico influenzano la ventilazione perché la CO2 reagendo con l’H2O porta alla formazione di HCO3e di H+. Quindi un aumento di CO2 si tramuterà in un aumento di H+, cioè in una diminuzione di pH • Le variazioni delle sostanze chimiche che sono percepite dai chemocettori riguardano quindi: • La CO2 • L’O2 • Gli ioni idrogeno (H+) • Una diminuzione della PO2 (ipossia), un aumento della PCO2 (ipercapnia) ed una diminuzione del pH del sangue arterioso sono tutti fattori che provocano una stimolazione della ventilazione alveolare • Se le variazioni sono di segno opposto vi sarà decremento della ventilazione I chemocettori • A seconda della loro localizzazione si distinguono in : • Periferici, localizzati nei glomi carotidei, vicino al seno carotideo, e nei glomi aortici, vicino all’arco dell’aorta • Centrali, localizzati nel midollo allungato Chemocettori periferici I Glomi carotidei: Piccoli organi pari localizzati nei pressi della biforcazione della catrotide comune, da cui ricevono l’apporto ematico Sono innervati da fibre pregangliari del simpatico ed inviano informazioni al n. del tratto solitario tramite fibre del nervo glossofaringeo Chemocettori periferici II • Glomi aortici: • Numerosi aggregati cellulari di varie dimensioni, arrangiati in quattro gruppi principali, irrorati da collaterali dell’aorta (sx) e della succlavia di destra (dx) • Sono innervati da fibre pregangliari del simpatico ed inviano informazioni al n. del tratto solitario tramite fibre del nervo vago Il glomo è costituito da accumuli di cellule glomiche (di tipo I) circondate da cellule di tipo II, simili alle cellule gliali. Le cellule glomiche sono reciprocamente connesse tramite gap junctions Le cellule glomiche sono a contatto, tramite il polo vascolare, con i capillari fenestrati, tramite il polo nervoso, con i dendriti dei nervi afferenti I glomi sono molto piccoli (circa 2 mg) hanno un indice metabolico molto elevato (9 ml di O2/min/100 gr), ed hanno il flusso ematico più elevato di tutto l’organismo (2 l/min/100 gr di tessuto) Le cellule glomiche sono ricche in canali ionici voltaggio-dipendenti In seguito a depolarizzazione, presentano potenziali d’azione Posseggono vescicole contenenti neurotrasmettitori (dopamina, acetilcolina, noradrenalina) che controllano la frequenza di scarica lungo le fibre afferenti Effetti dell’ipossia, ipercapnia, acidosi sui glomi Questi tre stimoli inducono nelle cellule una sequenza di eventi che comprende una inibizione dei canali del K, seguita da depolarizzazione della membrana e dall’apertura dei canali del Ca voltaggio-dipendenti che porta ad aumento della concentrazione intracellulare di Ca e a liberazione del neurotrasmettitore • In particolare, i meccanismi coinvolti, pur portando allo stesso effetto, sono di natura diversa: • Ipossia: i canali del K hanno un sensore (una proteina contenente eme) sensibile all’O2. In caso di ipossia, l’O2 non si lega al sensore, il canale resta chiuso, la permeabilità al K diminuisce, la cellula si depolarizza • Ipercapnia: agirebbe tramite l’ingresso di CO2 nella cellula con conseguente abbassamento di pH che determina inibizione dei canali K • Acidosi: stimolerebbe gli scambiatori acido-base (es. scambiatore Cl-HCO3-) con conseguente diminuzione del pH intracellulare e chiusura dei canali K Altri effetti che inducono attivazione glomica - Riduzione del flusso ematico attraverso i glomi - Caduta della P arteriosa - Aumento dell’attività del simpatico che induce vasocostrizione e quindi riduzione del flusso ematico Infatti, dato l’alto indice metabolico del tessuto, forti diminuzioni del flusso ematico inducono una caduta di PO2 a livello locale (anche se la P arteriosa dell’O2 rimane costante) con conseguente attivazione glomica Recettori chimici centrali • Questi recettori sono posti sulla superficie ventrolaterale del bulbo,subito caudalmente al ponte, in prossimità dei neuroni coinvolti nella ventilazione. Sono raggruppati in tre regioni: rostrale, intermedia e caudale • Le informazioni provenienti dalle regioni rostrali e caudali, convergono sulle regioni intermedie, dove vengono integrate con altre informazioni di natura vegetativa (P arteriosa, frequenza cardiaca, ecc).La distruzione dell’area intermedia blocca la chemocettività Stimoli adeguati per i recettori centrali • I recettori centrali rilevano la composizione del liquido cefalorachidiano. • In particolare, essi risponderebbero alle variazioni di concentrazione degli ioni H che a loro volta dipendono dalla PCO2 arteriosa e dalla concentrazione di ioni bicarbonato nello stesso liquor • Quindi essi risultano sensibili alle variazioni sia di PCO2 sia di pH ma NON alle variazioni di PO2 Attivazione dei chemocettori centrali Gli ioni H+ (es. derivati dall’ac. lattico) NON possono attraversare la barriera ematoencefalica. Invece, la CO2 diffonde dal sangue nel LCR Qui la CO2 non attiva direttamente i chemocettori ma, per la presenza dell’H2O, l’anidrasi carbonica catalizza la sua conversione in H2CO3 che si dissocia in ioni bicarbonato e ioni idrogeno Adesso gli H+ possono attivare i chemocettori centrali Risposta chemocettiva all’aumento di PCO2 Risposta chemocettiva all’aumento di H+ Sensibilità dei chemocettori centrali • I chemocettori centrali rendono conto del 75% dell’aumento della ventilazione indotto dall’aumento della CO2, quelli periferici solo del 25% • Un aumento della PCO2 arteriosa di 1mmHg, cioè del 2,5% (su 40mmHg) determina un incremento della ventilazione di 2l/min, cioè di circa il 40% Risposte dei chemocettori a variazioni prolungate delle pressioni parziali dei gas • I chemocettori centrali inizialmente rispondono intensamente alle variazioni della PCO2, stimolando la ventilazione, ma se la PCO2 permane elevata per più giorni, la ventilazione diminuisce • Infatti, la barriera ematoencefalica inizierà a trasportare nel liquor HCO3- che tamponeranno gli H+ e la stimolazione centrale diminuirà • Invece la risposta dei chemocettori periferici alle variazioni di O2, resta costante: quindi, in alcuni casi, la PO2 resta l’unico stimolo chimico per la ventilazione (es. pazienti con enfisema polmonare hanno ipossia e ipercapnia croniche, per cui, se si somministra loro troppo O2, potrebbero smettere di respirare, perché verrebbe a mancare l’unico stimolo chimico che supporta la ventilazione) Altri effetti che inducono attivazione centrale • Sembra che altri ioni possano influenzare la ventilazione (es. gli ioni potassio la stimolerebbero, gli ioni calcio la ridurrebbero) • La Temperatura può influenzare i chemocettori centrali direttamente o tramite i termocettori periferici (un improvviso raffreddamento del corpo stimola la inspirazione, mentre una immersione in acqua calda determina iperventilazione) • Aumento di Adrenalina e noradrenalina in circolo induce iperventilazione (esercizio fisico, stress, stati di eccitazione mentale) • Anche progesterone ed ormoni tiroidei determinano aumento della ventilazione….. Controllo chimico del respiro (sintesi) • I glomi sono influenzati dall’ipossia, dall’ipercapnia e dall’abbassamento del pH • I glomi controllano circa il 25% della ventilazione polmonare • I chemocettori bulbari rispondono a ipercapnia e conseguente abbassamento del pH • I chemocettori bulbari non sono stimolati dall’ipossia • I chemocettori controllano circa il 75% della ventilazione polmonare Regolazione nervosa del respiro • L’adeguamento della ventilazione alle esigenze metaboliche dell’organismo implica l’adeguamento della contrazione dei vari muscoli che controllano le variazioni volumetriche del torace • Sia i muscoli principali, sia i muscoli accessori sono innervati da motoneuroni situati nel MS a livello cervicale e toracico • Questi motoneuroni sono sottoposti a controllo da parte di centri sopraspinali che, a loro volta, risentono delle variazioni della PO2, PCO2 e della concentrazione di H+ • Al fine di individuare le strutture soprasegmentali implicate in tale controllo, in animali da esperimento sono state effettuate sezioni a diversi livelli del tronco encefalico Individuazione delle strutture implicate nel controllo nervoso del respiro Una sezione tra MS e bulbo (IV sezione), che interrompe tutte le connessioni nervose con i centri superiori, porta ad arresto del respiro e morte dell’animale. I II III Una sezione al di sopra del ponte (I sezione) non altera la IV IV respirazione: i centri superiori al ponte non sono necessari ma partecipano alla regolazione del respiro nel corso di attività Quindi il controllo del respiro è motorie volontarie più o meno a carico di strutture localizzate complesse quali canto, nel bulbo-ponte deglutizione, fonazione, tosse… Se in un animale integro si opera una resezione dei nervi vaghi al collo, il respiro rimane ritmico ma diventa più profondo e raro La seconda sezione (decerebrazione con rigidità degli arti) elimina la porzione ventrorostrale del ponte, in particolare i nn. parabrachiale mediale e di Kölliker-fuse, definiti centro pneumotassico: la respirazione risulta sostanzialmente inalterata Se si combina la vagotomia al collo con la decerebrazione, si assiste al fenomeno dell’apneusi: lunghe inspirazioni separate da corte espirazioni Operando una sezione più caudale, tra bulbo e ponte (III sezione), viene eliminato un centro posto nella reticolare pontina detto centro apneustico. L’apneusi scompare e si manifesta un respiro irregolare, spasmodico, detto gasping Deduzione • Da tutto questo si può dedurre che, poiché non scompare dopo resezione pontina, il ritmo respiratorio è a carico del bulbo (dove saranno localizzati i neuroni inspiratori ed espiratori), mentre frequenza e profondità sono a carico del ponte Relazioni esistenti tra i centri pontini Il fenomeno dell’apneusi è stato spiegato nel modo seguente: • Il centro pneumotassico svolge un’azione inibente sul centro apneustico (così come il vago, che riceve informazioni dai recettori di tensione polmonari) ed attivante sui neuroni espiratori bulbari • Il centro apneustico è un centro che attiva tonicamente i neuroni inspiratori bulbari • Venendo a mancare sull’apneustico la doppia inibizione del vago e dello pneumotassico, viene liberata la sua azione tonica attivante per cui i neuroni inspiratori scaricano più a lungo dando luogo a lunghe inspirazioni • D’altro canto i neuroni espiratori non sono più attivati dal centro pneumotassico e quindi le espirazioni sono di breve durata Schema dei rapporti esistenti tra i centri che controllano il respiro PN = centro pneumotassico AP = centro apneustico IN = neuroni inspiratori ES = neuroni espiratori MI = midollo cervicale • L’andamento normale della ventilazione dipende dalla comunicazione tra bulbo e ponte, ma il centro di controllo principale della ventilazione, quello che regola il ritmo respiratorio, è nel bulbo I centri bulbari Il centro respiratorio bulbare o centro generatore del ritmo respiratorio, e che comprende i “centri” inspiratorio ed espiratorio, non è costituito da un raggruppamento ben localizzato di neuroni come farebbe supporre la definizione n. inspiratori Si tratta, invece di una rete neuronale in cui i neuroni inspiratori ed espiratori sono mescolati ed interconnessi fra loro, anche se per comodità si continua a parlare di “centri” n. espiratori I centri bulbari • Le strutture che costituiscono il centro respiratorio sono: • Il Gruppo Respiratorio Ventrale (GRV), nei pressi del n. ambiguo, contenente neuroni sia inspiratori, sia espiratori • Il Gruppo Respiratorio Dorsale (GRD), nella regione del n. del tratto solitario, contenente neuroni prevalentemente inspiratori I centri bulbari • GRV e GRD dei due lati sono connessi fra loro • I due gruppi sarebbero messi in azione da un aggregato neuronale, posto lateralmente che agirebbe come un integratore di base del ritmo respiratorio • Esso riceverebbe diversi segnali regolatori: volontà, stato emotivo, attività muscolare, dolore, temperatura corporea, ritmo sonno-veglia, stimoli chimici • GRD e GRV, da un punto di vista tonico, indipendentemente dal ritmo respiratorio, sono attivati dalla formazione reticolare che, a sua volta, riceve sia dalla periferia sia dai centri encefalici superiori • Le vie respiratorie sono crociate, quindi ciascun centro controlla i muscoli del lato opposto Classificazione dei neuroni bulbari • La classificazione dei neuroni respiratori bulbari localizzati nei GRD e GRV comprende: • Neuroni propriobulbari: i cui assoni, brevi, non escono dal bulbo • Neuroni bulbospinali: i cui assoni raggiungono nel MS i motoneuroni dei muscoli respiratori • Neuroni attivati da recettori di stiramento polmonare: localizzati nel GRD insieme a neuroni propriobulbari Caratteristiche dei neuroni propriobulbari • N. inspiratori precoci: (GRV intermedio) si attivano all’inizio dell’inspirazione (I) e si spengono verso la fine • N. inspiratori o della rampa inspiratoria: (GRV intermedio) scaricano per tutta l’I con aumento graduale della frequenza. Attivano i n. bulbospinali inspiratori • N. inspiratori tardivi: (GRD) scaricano alla fine dell’I • N. postinspiratori: (GRD)scaricano poco dopo la fine dell’I • N. espiratori (tardivi): (GRD) scaricano durante l’espirazione (E) • N. preinspiratori: (GRV rostrale) scaricano alla fine dell’E e all’inizio dell’I (non mostrati in figura) Caratteristiche dei neuroni bulbospinali • N. inspiratori bulbospinali: (GRV intermedio) inviano segnali al MS in stretto andamento con la scarica dei n. inspiratori della rampa • N. espiratori bulbospinali: (GRV caudale) inviano segnali al MS nella seconda fase dell’E • N. inspiratori ed espiratori per la respirazione forzata: (GRV caudale) sono alcuni dei neuroni già descritti che entrano in azione solo durante la respirazione forzata, per attivare i muscoli respiratori accessori Caratteristiche dei neuroni attivati dai recettori da stiramento polmonare • Neuroni Rα ed Rβ: i primi inibiti, i secondi attivati dall’insufflazione del polmone • Neuroni P (pompa): attivati dai recettori di stiramento polmonare e in rapporto con le variazioni di volume polmonare • Sono implicati nel riflesso di Hering-Breuer Riassumendo: • Nel GRD (n. del tratto solitario) troviamo i neuroni del gruppo inspiratorio: - Rα, Rβ, P, inspiratori tardivi, postinspiratori • Nel GRV, diviso in porzione rostrale, intermedia e caudale, troviamo neuroni misti, inspiratori ed espiratori: - Porzione rostrale: n. espiratori propriobulbari - Porzione intermedia (n. paraambiguale): n. inspiratori propriobulbari (inspiratori precoci e a rampa) e n. bulbospinali che si portano ai neuroni del nervo frenico, degli intercostali esterni e dei mm. accessori contralaterali - Porzione caudale (n. retroambiguale): n. espiratori bulbospinali che vanno ai neuroni dei mm. addominali ed intercostali interni Sequenza di attivazione dei neuroni respiratori 1) Stimoli periferici e centrali attivano i nn. propriobulbari inspiratori precoci e della rampa. La velocità di salita della rampa dipende dal flusso degli impulsi che stimolano i nn. 2) I neuroni della rampa attivano i neuroni bulbospinali inspiratori che, a loro volta, attivano i muscoli inspiratori: espansione della cassa toracica (c.a 2 sec) Entrano in azione i neuroni inspiratori tardivi, i postinspiratori, gli espiratori precoci e si ha inibizione graduale dei neuroni propriobulbari e bulbospinali Ha inizio l’espirazione, fenomeno passivo dovuto al ritorno elastico dei polmoni (c.a 3 sec). Tuttavia nella II fase dell’espirazione entrano in azione gli espiratori tardivi che attivano i bulbospinali espiratori (forse per rallentare il flusso aereo in uscita). Gli espiratori tardivi inibiscono gli inspiratori, tranne i precoci che ricominciano a scaricare verso la fine dell’espirazione (gli inspiratori vengono inibiti anche dalle influenze dei recettori da stiramento polmonare) 3) 4) 5) Sequenza respiratoria 1) Attivazione inspirazione 2) Generazione del pattern inspiratorio 3) Inattivazione dell’inspirazione 4) Attivazione dell’espirazione 5) Inattivazione dell’espirazione Il centro respiratorio: automatico o riflesso? • Il ritmo è una proprietà intrinseca del centro respiratorio o è mantenuto da impulsi afferenti dai recettori? • La maggior parte degli studiosi ritiene il centro respiratorio bulbare un centro automatico • Però è fuor di dubbio che la sua attività può essere modulata da impulsi afferenti di varia natura e quindi esso funziona anche da centro riflesso Centro automatico • Si ritiene che la rete nervosa responsabile del ritmo respiratorio consti di due parti: - un pace-maker (vero generatore del ritmo endogeno) - una parte da cui nascono gli impulsi codificati dell’attività dei premotoneuroni • Il pace-maker sarebbe localizzato davanti alla porzione rostrale del GRV (complesso preBöttzinger, tra il n. ambiguo e il n. reticolare laterale) e scaricherebbe ritmicamente grazie alle sue caratteristiche intrinseche di membrana • Esso invierebbe impulsi ad alcune regioni troncoencefaliche (GRV, ad es.) contenenti premotoneuroni (connessi ai motoneuroni) che trasformerebbero l’uscita ritmica dell’oscillatore in un complesso di segnali organizzati nello spazio e nel tempo Centro riflesso • I centri respiratori possono funzionare anche da centri riflessi. • I riflessi possono essere di origine: - polmonare, a partenza da recettori da stiramento (Hering-Breuer), intraepiteliali (stimolati da sostanze irritanti), alvelolari nocicettivi (J-recettori, sensibili alla congestione polmonare e ai microembolismi) - nasale (sternuto) - dall’epifaringe (da aspirazione) - laringei e tracheali (tosse) Riflesso di Hering-Breuer • La distensione dei polmoni determina in via riflessa un atto espiratorio, mentre la loro desufflazione ne provoca uno inspiratorio • Riflesso di origine polmonare: nell’Uomo ha importanza ridotta, ma entra in gioco quando la compliance (distensibilità) polmonare è ridotta (congestione polmonare, edema polmonare…) oppure per impedire la sovradistensione polmonare sotto sforzo (se il Vol corrente supera 1lt, contro i normali 500 ml) Riflesso di Hering-Breuer • A seguito di sovradistensione polmonare o di eccessiva insufflazione vengono stimolati i tensocettori polmonari ed i recettori da irritazione • Le afferenze di questi recettori, tramite fibre vagali, penetrano nel bulbo e terminano nel GRD andando ad inibire i neuroni Rα e ad attivare gli Rβ: ciò determina inibizione della inspirazione con conseguente atto espiratorio riflesso. • La desufflazione ha l’effetto opposto: i tensocettori non vengono stimolati, non inibiscono il centro respiratorio e si ha l’inspirazione • Il riflesso scompare a seguito di resezione vagale Lo sternuto • Riflesso di origine nasale: stimolazioni a livello delle narici o delle cavità nasali generano un riflesso protettivo che consiste in una inspirazione seguita da una espirazione piuttosto rumorosa. • La via afferente è rappresentata da rami del nervo trigemino che hanno la cellula nel ganglio di Gasser • La via efferente interessa non solo il midollo per i muscoli respiratori, ma anche il nervo facciale per la muscolatura mimica Riflesso da aspirazione • Il riflesso ha origine nell’epifaringe (o rinofaringe): riflesso protettivo, consistente in una rapida inspirazione grazie alla quale il materiale viene sospinto nella sottostante faringe per ripulire le vie nasali • La via afferente è rappresentata dal ramo faringeo del nervo glossofaringeo La tosse Riflesso protettivo: ha origine per irritazione o processi patologici che possono interessare laringe, trachea, bronchi, polmoni, pleure - Le vie afferenti sono rappresentate dal vago (nervo laringeo superiore) e dal nervo glossofaringeo - Le vie efferenti sono rappresentate da impulsi che scendono lungo il midollo per i mm. respiratori e lungo il vago (nervi laringei), per i mm. della laringe - Si verifica un atto inspiratorio profondo seguito da chiusura della glottide e da atti espiratori corti e più o meno violenti eseguiti a glottide chiusa, allo scopo di allontanare gli agenti irritanti - Durante la tosse la pressione intrapleurica e dentro i bronchi diventa positiva e può raggiungere i 300 mmHg - Quando si apre la glottide, forzata dall’atto inspiratorio (essa resta chiusa 0,2 sec), il flusso d’aria può arrivare a 6 lt/sec con una velocità prossima a quella del suono Sonno e ventilazione • Durante il sonno non-REM, la ventilazione viene regolata esclusivamente dalle pressioni parziali dei gas respiratori • Ciò accade perché diminuisce l’attività del sistema reticolare troncoencefalico che è coinvolto sia nello stato di allerta del SNC, sia nella regolazione della ventilazione (eccitazione tonica dei neuroni respiratori midollari) • In concomitanza con gli eventi fasici del sonno REM si assiste ad un aumento di ventilazione • In generale il sonno determina una depressione dell’attività respiratoria Il primo atto respiratorio Prima della nascita la P intrapleurica non è al di sotto di quella atmosferica Alla nascita, la legatura del cordone ombelicale induce un drammatico aumento della PCO2 e diminuzione della PO2 nel sangue del neonato Vengono così stimolati i chemocettori centrali e periferici ed insorge il primo atto inspiratorio Nel passaggio nel canale genitale materno, il feto ha espulso parte dei liquidi contenuti nel polmone per compressione del torace, ma le prime vie aeree contengono ancora liquido con una viscosità maggiore dell’aria Nell’entrare nell’apparato respiratorio l’aria dovrà superare una notevole resistenza e la P intrapleurica diventerà molto negativa (-50/60 cm H2O) Segue la prima espirazione, in cui la P intrapleurica è alta (+ 20 cmH20) Il liquido che residua nel polmone viene riassorbito tramite i capillari ematici e linfatici In seguito si forma il surfactante che riduce la tensione superficiale del film liquido che bagna gli alveoli e le resistenze diminuiscono Altre modificazioni del respiro • Il respiro può essere modificato anche per cause volontarie o involontarie • Tra le modificazioni del respiro ricordiamo: - il singhiozzo - lo sbadiglio - il pianto - il riso Il singhiozzo Consegue all’irritazione del nervo frenico che si contrae ritmicamente. Ogni contrazione del diaframma si conclude con una brusca chiusura della glottide, determinando il tipico suono “hic”. Il motivo scatenante il singhiozzo non si conosce, ma alcune situazioni tipiche della vita quotidiana, lo possono indurre: - dilatazione dello stomaco, determinata dalla rapida o eccessiva ingestione di cibo e liquidi - bruschi sbalzi di temperatura, come bere una bevanda bollente o gelata - l’eccessiva ingestione di bevande alcoliche può infiammare la mucosa gastrica e, indirettamente, irritare il diaframma - episodi di emotività: in condizione di forte disagio, si ingoia una quantità di aria superiore al normale; ciò provoca come diretta conseguenza l'irritazione del diaframma e quindi la comparsa del singhiozzo. Lo sbadiglio • E’ dovuto ad una inspirazione lenta e profonda con apertura forzata della bocca, delle fauci e della glottide; viene considerato un atto respiratorio accessorio. • Può essere volontario o determinato da fattori organici (fame, sonno) e psichici (noia, stanchezza, astenia ecc.) e può essere modulato Effetti dello sbadiglio • Lo sbadiglio permette di prendere una grande quantità di ossigeno, stimolare l'attività cellulare (così da produrre energia da usare come carburante, per compensare una carenza energetica), attivare la circolazione e rilassare i muscoli, scaricando tensione • Sbadigliare aiuta anche a lenire il dolore: l'aumento dell'ossigeno nel sangue spegne la mente "razionale", ci si distrae e il nostro organismo usa le energie risparmiate per produrre sostanze che calmano il dolore. L'inibizione della mente razionale libera le capacità più intuitive e creative e aumenta le capacità percettive • Lo sbadiglio è contagioso: probabilmente si tratta soltanto di un comportamento d'imitazione. Il pianto • Si tratta di una reazione a dolore o emozione • Consiste in una serie di inspirazioni ripetute ed espirazioni prolungate, accompagnate da suoni e da iperlacrimazione • Nella manifestazione del pianto vengono coinvolti: secrezione lacrimale, atteggiamento emozionale del viso, vocalizzazioni e lamenti • Il pianto produce enkefalina, un oppioide endogeno e potente anestetico la cui funzione principale è di scaricare la tensione accumulata, rilassando i muscoli. • Un'altra funzione è quella di lavare la cornea (tramite l’incremento della produzione del liquido lacrimale) nel momento in cui ci sia un corpo estraneo che la irrita. Il riso • Si fa una profonda inspirazione seguita da una profonda espirazione accompagnata da un suono irregolare: i muscoli del volto, del collo, del diaframma, dell'addome sono in tensione. Quando il fiato è esaurito, si inspira altra aria a i suoni riprendono. Il riso può essere accompagnato da lacrimazione • La risata coinvolge entrambi gli emisferi cerebrali. Essa attiva una zona corticale che assiste l'area motoria principale nel controllo dei movimenti. • Ridere è un'attività energetica che aumenta il battito cardiaco e la pressione del sangue • La risata è una sorta di linguaggio sociale che permette di mettersi in relazione con gli altri • La risata è una caratteristica innata che l'uomo condivide solo con i suoi cugini primati, gli scimpanzé, i gorilla e le altre scimmie.
Scaricare