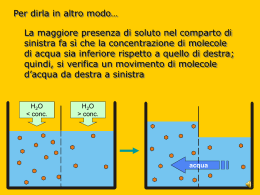
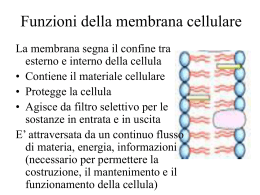
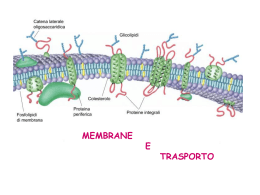
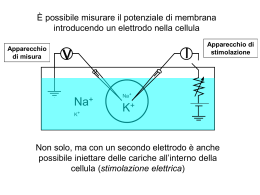
Trasporti attraverso la membrana Proteine di trasporto Una membrana costituita da un puro bilayer fosfolipidico è impermeabile alle proteine e alla maggior parte delle piccole molecole Gas Piccole molecole polari non cariche Etanolo Acqua Urea Grosse molecole polari non cariche Glucosio Ioni Molecole polari cariche Aminoacidi ATP Glc-6-P Proteine di trasporto 2 principali classi di proteine di trasporto: Proteine Carrier Proteine Canale soluto ione bilayer lipidico sito di legame del soluto poro acquoso Diffusione Facilitata Caratteristiche dei trasporti mediati • I carriers sono dotati di specificità • Sono soggetti a saturazione • Possono essere bloccati dagli inibitori competitivi • Hanno un’elevata dipendenza termica e dal pH I trasportatori hanno le caratteristiche di enzimi • I carriers agiscono cataliticamente come gli enzimi • Legano selettivamente il loro substrato, cioè la molecola che deve essere trasportata • Cambiano di conformazione per rilasciare il substrato dall’altro lato • Ritornano alla conformazione originale per legare un’altra molecola di substrato • Seguono una cinetica del tipo Michaelis-Menten Analisi cinetica del transporto di una molecola tramite proteina carrier: saturazione In base alla Ia legge di Fick il flusso di particelle che diffondono liberamente aumenta linearmente all’aumentare della concentrazione Ma la Ia legge di Fick non viene più rispettata se si tratta di un flusso di particelle attraverso la membrana mediato da carriers 20 300 15 F=kdDC 200 F F 2 Flusso netto [moli/(cm s] Fmax 10 100 F max ka 1 C I flussi mediati da carriers a differenza della diffusione libera - sono saturanti 0 5 0 10 DC 20 0 0 50 100 150 30 200 104 DC (mM) Ciò accade per due motivi: 1. Sulla membrana è presente un numero finito di carriers; 2. Ciascun carrier opera ad una velocità finita Rappresentazione del concetto di saturazione con un esempio numerico Per semplicità consideriamo una membrana con un solo carrier Velocità del carrier: 50 p/s Flusso 1 10 50 50 50 50 Flusso (Particelle/Carrier/s) N. partic. 1 10 50 100 1000 40 30 20 10 0 0 800 1000 N. particelle Quesito su cui meditare Distruggendo la metà dei carriers sulla membrana, Fmax rimarrebbe inalterata, aumenterebbe o diminuirebbe? Come è stato ottenuto il grafico che illustra come varia il flusso al variare della concentrazione? 2 Flusso netto [moli/(cm s] 20 C1 15 ? 10 5 F1 Cellule in sospensione 0 0 20 40 60 80 DC (mM) pendenza della retta Si introduce nella provetta substrato S radioattivo ad una concentrazione C1 2. A tempi successivi (t0, t1, t2, t3, …) si preleva un campione dalla provetta e si misura la concentrazione di S radioattivo all’interno delle cellule del campione 3. Normalizzando la velocità v1 rispetto all’area della membrana si ottiene il primo valore del flusso F1 riferito alla concentrazione C1 100 C1 5 1. D[ S ]in v1 Dt 4 [S]in 3 2 1 0 0 10 20 30 Tempo 40 50 2 Flusso netto [moli/(cm s] 20 F4 F3 15 Successivamente si introduce in ciascuna provetta substrato S radioattivo alle altre concentrazioni C2, C3, C4 …. crescenti e si ripete la stessa procedura descritta precedentemente F2 10 5 F1 C2 0 0 C2 20 C3 40 60 DC (mM) 80 C4 100 C1 V4F4 40 30 V3F3 [S]in 20 V2F2 10 0 0 10 20 Tempo 30 40 50 C3 C4 I carriers, come gli enzimi, possono essere soggetti ad inibizione competitiva 2 Flusso netto [moli/(cm s] Fmax 20 15 +Ic 10 +Ic 5 0 0 50 100 150 200 104 DC (mM) Quindi, in presenza di inibitore competitivo il valore di Fmax non cambia. In presenza di Ic varia ka Calcolo della costante di affinità ka F max 20 2 Flusso netto [moli/(cm s] 2 Flusso netto [moli/(cms] 20 15 10 5 0 15 5 0 0 50 100 DC (mM) 150 200 F max 2 10 ka3 0 ka1 ka2 10 DC (mM) 20 30 ka è quel valore di concentrazione del substrato al quale il flusso è la metà di quello massimo ka è inversamente proporzionale all’affinità del carrier per il substrato Come funziona un inibitore competitivo? [S] [Ic] Prob .01 10 .001 .1 10 .010 1 10 .091 10 10 .5 100 10 .909 1000 10 .990 10000 10 .999 solo S S << Ic S >> Ic substrato Inibitore comp. 20 Se 15 si vuole costruire un grafico che rappresenti Sun range di 10 S+Ic concentrazioni molto ampio (alcuni 5 ordini di grandezza) conviene usare una scala logaritmica 0 0 2000 4000 6000 [S] 8000 10000 15 Flusso Flusso 20 10 5 S S+Ic 0 0.01 0.1 1 10 [S] 100 1000 10000 Quesito del giorno Un ricercatore trova che la velocità con cui una sostanza è trasportata all’interno di certe cellule varia al variare della sua concentrazione come illustrato in tabella. 1. Trovare i corrispondenti valori di flusso sapendo che l’area di membrana su cui sono state fatte le misure è 3·10-2 cm2; 2. Rappresentare graficamente i valori del flusso al variare della concentrazione di substrato in due grafici distinti ove le concentrazioni sono rappresentate rispettivamente in forma lineare e logaritmica; 3. Ricavare dal grafico i valori di Fmax e ka. Conc. mM v (mmol/s) 0.1 3.0 1 10.0 5 16.7 10 18.2 20 19.0 30 19.4 50 19.6 100 19.8 200 19.9 Risposta al quesito 600 600 500 500 mmoli/s/cm mmoli/s/cm 2 700 2 700 400 300 200 100 400 300 200 100 0 0 50 100 Concentr. 150 200 0 0.01 0.1 1 Concentr. 10 100 Esempio di Diffusione facilitata Il Trasportatore del Glucosio Glucosio Permeasi extracell. intracell. poro: formato da 5 a-eliche transmembrana anfipatiche Diffusione Facilitata soluto stato B stato A esterno bilayer lipidico gradiente di concentrazione interno proteina carrier sito di legame del soluto es. Trasportatore del glucosio negli epatociti La driving force per il tranporto in qualsivoglia direzione è determinata dal gradiente di concentrazione attraverso la membrana Attivita' di Ricerca Responsabili Prof. M. Toselli e Dr. Gerardo Biella Il LABORATORIO DI FISIOLOGIA E BIOFISICA DEI CANALI IONICI del Dipartimento di Fisiologia e Farmacologia Cellulari e Molecolari dell'Universita' di Pavia, diretto dal Prof. Mauro Toselli svolge attivita’ di ricerca sui seguenti argomenti: l Canali ionici voltaggio-dipendenti: proprieta' biofisiche e loro correlazione ad aspetti funzionali dell'eccitabilita' elettrica nei neuroni durante lo sviluppo. l Modulazione dei canali del calcio e Regolazione del traffico intracellulare: meccanismi molecolari e biofisici dei processi di regolazione dell'attivita' dei canali del calcio voltaggio-dipendenti da parte di protein chinasi e proteine G, e conseguenze nel controllo della trasmissione, ricezione e immagazinaggio dell'informazione nei neuroni. l Neurofisiologia cellulare: Identificazione e caratterizzazione funzionale dei circuiti sinaptici delle cortecce paraippocampali. In particolare: ruolo che l'attività ritmica sincronizzata dei neuroni del sistema limbico esercita sui processi di apprendimento e memorizzazione. l Cellule staminali neuronali: caratterizzazione elettrofisiologica di cellule staminali neuronali in coltura. Le tecniche correntemente impiegate comprendono misurazioni elettrofisiologiche di patch-clamp sui seguenti preparati: a) linee cellulari stabili di origine neuronale, wild-type o opportunamente ingenierizzate per l'espressione di specifiche proteine ricombinanti normali o mutagenizzate; b) colture primarie di neuroni; c) sezioni sottili di tessuto; d) cellule staminali in coltura. Numero di posti disponibili: tre posti disponibili dall’inizio del secondo semestre a dicembre 2005; Requisiti per l'accettazione: verra’ data la priorita’ agli studenti che abbiano superato gli esami di Fisica e Chimica Generale. Alcune considerazioni di bioenergetica L’energia è la capacità di compiere un lavoro Energia del Sistema Un sistema all’equilibrio non può compiere un lavoro. Esso è al minimo della collina energetica Equilibrio La parte di Energia che il sistema può utilizzare per compiere un lavoro è definita energia libera del sistema Le reazioni non all’equilibrio spontaneamente procederanno nella direzione che porta all’equilibrio Tali reazioni sono dette esoergoniche Una reazione può essere forzata a procedere in senso non spontaneo. In questo caso la reazione assorbe energia: cioè occorre compiere un lavoro sul sistema Una reazione di questo tipo è detta endoergonica Le reazioni chimiche accoppiate Un processo endoergonico può avvenire se esso viene opportunamente accoppiato ad un processo esoergonico Consideriamo la seguente reazione endoergonica: A + B A-B DG>0 Affinchè essa possa avvenire deve attingere energia da una reazione esoergonica: X-P X + P + Energia DG<0 La sequenza di reazioni accoppiate sarà la seguente: A + X-P A-P + X A-P + B A-B + P I contenitori di energia nelle cellule Legami ad alta energia: ATP GTP Gradienti transmembranari: Gradienti ionici a cavallo del plasmalemma Contenitori intracellulari di Ca2+ Trasporti passivi e attivi a confronto molecola trasportata proteina carrier gradiente di concentraz. bilayer lipidico diffusione semplice mediata da canale mediata da carrier trasporto passivo trasporto attivo Trasporti passivi e attivi a confronto • Il gradiente di concentrazione attraverso la membrana determina la direzione e la velocità della diffusione passiva. • Nella diffusione passiva le molecole si muovono dalla zona ad alta concentrazione a quella a bassa concentrazione (trasporto “in discesa”) • Diffusione semplice: le molecole diffondono attraverso il bilayer lipidico • Diffusione facilitata: diffusione attraverso la membrana mediata da proteine di trasporto (non è richiesta energia) • Muovere un soluto “in salita” (contro il gradiente di concentrazione) richiede un ingresso di energia Trasporto Attivo Trasporti Attivi Il trasporto di un soluto contro un gradiente elettrochimico richiede l’utilizzo di energia Due strategie sono adottate dalle cellule animali: Pompe ATP-dipendenti Trasporti accoppiati Gradiente elettrochimico Trasporto in salita accoppiato direttamente all’idrolisi dell’ATP Trasporto in salita di una molecola accoppiato al trasporto in discesa di un’altra Na+/K+ ATPasi Na+/K+ ATPasi sito di legame per K+ e ouabaina gradiente di K+ gradiente di Na+ cytosol sito di legame del Na+ Na+/K+ ATPasi • La pompa idrolizza l’ATP ad ADP per trasportare simultaneamente 3 Na+ dalla cellula e 2 K+ dentro la cellula ad ogni ciclo della pompa • La Na+-K+ ATPasi è responsabile di >30% del consumo totale di ATP • Mantiene un gradiente del Na+ (ext>int) e del K+ (int>ext) • Contribuisce a creare un potenziale di membrana negativo all’interno • È bloccata dalla ouabaina Ione Intracellulare Na+ K+ 5-15 mM 140 mM Extracellulare 145 mM 5 mM Ciclo della pompa Na+/K+ ATPasi la proteina subisce un cambiamento conformazionale, il Na è rilasciato all’esterno fosforilazione ATP-dipendente ext int il Na si lega al sito citosolico il K si lega al sito extracellulare la proteina ritorna alla conformazione originale, il K è rilasciato all’interno defosforilazione 1 ciclo 10 millisecondi Alcune importanti caratteristiche della pompa Velocità di attività dipendente dalla concentrazione intracellulare di Na+ Bloccata dal glicoside cardiaco ouabaina e bassa [ATP]i Stechiometria - 3 Na+ fuori, 2 K+ dentro, 1 ATP usato Elettrogenica, può generare direttamente da -2 a -20 mV Tiene la pressione osmotica sotto controllo, prevenendo il rigonfiamento cellulare Gradiente elettrochimico • A cavallo della membrana della maggior parte delle cellule c’è una differenza di potenziale elettrico – il potenziale di membrana • Il potenziale di membrana influenza il movimento transmembrana di tutte le molecole cariche (ioni) • Il lato citoplasmatico della membrana plasmatica di solito ha un potenziale più negativo rispetto all’esterno • La forza elettrostatica spinge i cationi nella cellula e guida gli anioni fuori • Quindi, quando consideriamo la diffusione passiva di soluti carichi attraverso una membrana, due forze devono essere considerate: • (a) il gradiente di concentrazione transmembrana • (b) la differenza di potenziale transmembrana • La driving force netta = Gradiente Elettrochimico Gradiente Elettrochimico fuori dentro potenziale di membrana=0 potenziale di membrana negativo dentro trasporto dei cationi aumentato potenziale di membrana positivo dentro trasporto dei cationi ridotto Quesito del giorno In base a quanto detto a lezione, stabilire se la pompa Na+/K+, che è elettrogenica dal momento che trasporta un numero netto di cariche elettriche, aumenterà o diminuirà la sua velocità d’azione qualora la membrana venga depolarizzata. Ricordarsi che una depolarizzazione comporta un aumento delle cariche positive all’interno della cellula rispetto alla situazione di riposo (potenziale di riposo). Alcune definizioni molecola trasportata ione co-transportato ione controtransportato Uniporto Simporto Antiporto Trasporti accoppiati Altri esempi di trasporti attivi primari Trasporti accoppiati Il gradiente di un soluto (es. gradiente del Na+) viene usato per guidare in salita il trasporto di una seconda molecola - trasporto attivo secondario Gradiente elettrochimico Trasporto in salita di una molecola accoppiato al trasporto in discesa di un’altra Trasporti accoppiati al gradiente di Na Nelle cellule animali molti processi di trasporto di membrana sono accoppiati al gradiente del Na. Es. Trasporto del glucosio in cellule dell’epitelio intestinale Na+ gradiente di Na carrier stato stato A B glucosio ext gradiente di glucosio int Stechiometria: 2 Na+ 1 Glc Bloccante: florizina Altri esempi di trasporti attivi secondari Trasporto del glucosio dal lume intestinale al sangue Sangue GLU permeasi Glucosio nel sangue Canale del K+ Cellule dell’epitelio intestinale Glucosio assunto Co-trasportatore Na/GLU Lume intest. Quesito del giorno Una cellula si trova immersa in una soluzione extracellulare contenente glucosio 10 mM. Trascorso un certo tempo la concentrazione di glucosio nel citoplasma sale a 50 mM. Secondo voi, perche’ cio’ prova che la membrana plasmatica di quella cellula dispone di un trasportatore attivo del glucosio? Come fareste a dimostrare sperimentalmente che tale trasportatore e’ Na+-dipendente (trasporto attivo secondario)? Mantenimento dei livelli di Ca2+ intracellulare • Il Ca2+ è coinvolto in molti processi cellulari • Il mantenimento di bassi livelli di Ca2+ intracellulare è critico for il normale funczionamento cellulare • La [Ca2+] extracellulare >>>> della [Ca2+] intracellulare • Le pompe del Ca2+ ATP-dipendenti della membrana plasmatica e del reticolo endoplasmatico pompano attivamente Ca2+ fuori dal citoplasma • Esiste anche uno scambiatore Na/Ca (T.A.II) che pompa attivamente Ca2+ fuori dalla cellula Quali meccanismi mantengono l’ampio gradiente di calcio? 3Na + ESTERNO INTERNO ATP Ca++ ADP Pompa del Calcio Ca++ Scambiatore Sodio:Calcio I gradienti di calcio esistono sia all’interno della cellula che a cavallo della membrana plasmatica Contrazione Ca2+ Secrezione Ca2+ + Esocitosi Ca2+ Ca2+ Espressione di geni Secondo messaggero Ca2+ Ca2+ Na+ Concentrazioni ioniche intracellulari ed extracellulari Ione Intracellulare Extracellulare Na+ K+ Mg2+ Ca2+ H+ 5-15 mM 140 mM 0.5 mM 10-7 mM 10-7.2 M (pH 7.2) 145 mM 5 mM 1-2 mM 1-2 mM 10-7.4 M (pH 7.4) ClAnioni fissi 5-15 mM high 110 mM 0 mM La [intracellulare] è molto diversa dalla [extracellulare] I cationi sono bilanciati dagli anioni C:\Documents and Settings\Administrator\Documenti\Didattica\Univ British Columbia Biology 350 Bioenergetics.doc
Scaricare