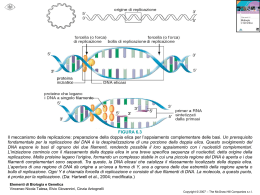
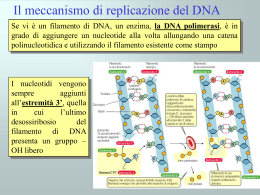
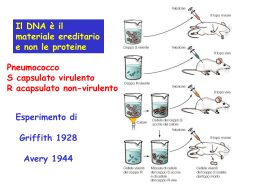
ALLA SCOPERTA DEL DNA STORIA Miescher 1869: il biochimico svizzero Friedrich Miescher individua una sostanza microscopica contenuta nel pus di bende chirurgiche utilizzate. Dal momento che tale molecola aveva la sua localizzazione nel nucleo, egli la chiamò nucleina. 1919:Phoebus Levene individuò la struttura del nucleotide, composta da base azotata, zucchero e fosfato. suggerì che il DNA consistesse in un filamento di nucleotidi legati tra loro attraverso i fosfati. Griffith 1928:Frederick Griffith scoprì, che i caratteri della forma Levene smooth, (liscia) di Pneumococcus potevano essere trasferiti alla forma rough (rugosa) miscelando i resti di batteri smooth morti con batteri rough vivi. Questo sistema, pur non fornendo nessuna evidenza su quale fosse la sostanza che determinava il cambiamento, mostrava comunque che qualcosa potesse trasportare l'informazione genetica dai resti dei batteri morti a quelli vivi. Si parlò quindi di un principio trasformante in grado di modificare i batteri vivi Hershey e chase 1937 :William Astbury presentò i primi risultati di alcuni studi di diffrazione a raggi X, che dimostrarono che il DNA ha una struttura estremamente regolare. Avery 1943 :Oswald Theodore Avery dimostrò in un celebre esperimento insieme a Colin MacLeod e Maclyn McCarty, che il DNA è il principio trasformante alla base di questo fenomeno. 1953:Alfred Hershey e Martha Chase dimostrarono attraverso un altro classico esperimento, che il materiale genetico del fago T2 è effettivamente il DNA. Astbury Nello stesso anno,attraverso ulteriori immagini da diffrazione a raggi X realizzate da Rosalind Franklin, chimica-fisica inglese, James Watson e Francis Crick presentarono sulla rivista Nature quello che è oggi accertato come il primo modello accurato della struttura del DNA,quello della doppia elica. A disegnarne il bozzetto fu Odile Speed, pittrice e moglie di Crick. Holley,Khorana e Nirenberg Watson e Crick 1957:In una importante presentazione Crick propose il dogma centrale della biologia molecolare, che fissa le relazioni tra DNA, RNA e proteine. 1958: La conferma finale del meccanismo di replicazione basato sulla struttura a doppia elica fu fornita dall'esperimento di Meselson-Stahl. Un successivo lavoro di Crick dimostrò come il codice genetico fosse basato su triplette di basi non sovrapposte, permettendo ad Har Gobind Khorana, Robert Holley e Marshall Warren Nirenberg di decifrarlo.Queste scoperte sono alla base della moderna biologia molecolare. Franklin 1962:dopo la morte di Rosalind Franklin (a causa di un tumore provocato, probabilmente, dalle alte dosi di raggi X a cui si era esposta nel corso dei suoi esperimenti), Watson, Crick e Wilkins ricevettero congiuntamente il Premio Nobel per la medicina COMPOSIZIONE Il DNA è un lungo polimero costituito da unità ripetute di nucleotidi. La catena del DNA è larga tra i 22 ed i 26 Ångström (da 2,2 a 2,6 nanometri) ed ogni unità nucleotidica è lunga 3,3 Ångstrom (0,33 nanometri).Sebbene ogni unità occupi uno spazio decisamente ridotto, la lunghezza dei polimeri di DNA può essere sorprendentemente elevata, dal momento che ogni filamento può contenere diversi milioni di nucleotidi. Ad esempio, il più grande cromosoma umano (il cromosoma 1) contiene quasi 250 milioni di paia di basi. Negli organismi viventi, il DNA non è quasi mai presente sotto forma di singolo filamento, ma come una coppia di filamenti saldamente associati tra loro.Essi si intrecciano tra loro a formare una struttura definita doppia elica. Ogni nucleotide è costituito da uno scheletro laterale, che ne permette il legame covalente con i nucleotidi adiacenti, e da una base azotata, che instaura legami idrogeno con la corrispondente base azotata presente sul filamento opposto. Il composto formato da una base azotata legata allo zucchero è definito nucleoside; un nucleotide è invece un nucleoside a cui sono legati uno o più gruppi fosfato. La struttura laterale del DNA è composta da unità ripetute ed alternate di gruppi fosfato e di 2-deossiribosio,uno zucchero pentoso (a cinque atomi di carbonio) che si lega ai fosfati adiacenti attraverso legami fosfodiesterici presso il terzo ed il quinto carbonio; Conseguenza di questi legami asimmetrici è che ogni filamento di DNA ha un senso, determinato dalla direzione dei legami fosfodiesterici. In una doppia elica, il senso di un filamento è opposto a quello del filamento complementare. Per tale motivo, i due filamenti che costituiscono una doppia elica sono detti antiparalleli. La principale differenza tra il DNA e l'RNA è lo zucchero pentoso utilizzato: l'RNA utilizza, infatti, il ribosio. La doppia elica del DNA è stabilizzata dai legami idrogeno che si instaurano tra le basi azotate presenti sui due filamenti. Le quattro basi che sono state individuate nel DNA sono l'adenina (abbreviata con la lettera A), la citosina (C), la guanina (G) e la timina (T). Adenina e guanina sono composti eterociclici chiamati purine, mentre citosina e timina sono anelli pirimidinici. Esiste una quinta base, di tipo pirimidinico, chiamata uracile (U), ma essa non è di norma presente nelle catene di DNA. L'uracile è altresì presente nei filamenti di RNA al posto della timina, da cui si differenzia per la mancanza di un gruppo metile. La doppia elica è una spirale destrorsa. Con l'avvitarsi su sé stessi dei due filamenti, restano esposti dei solchi tra i diversi gruppi fosfato. APPAIAMENTO DELLE BASI Ogni tipo di base presente su un filamento forma un legame con la base posta sul filamento opposto. Tale evento è noto come appaiamento complementare. Le basi puriniche formano legami idrogeno con le basi pirimidiniche: A può legare solo T e G può legare solo C. L'associazione di due basi viene comunemente chiamata paio di basi ed è l'unità di misura maggiormente utilizzata per definire la lunghezza di una molecola di DNA. Dal momento che i legami idrogeno non sono covalenti, essi possono esser rotti e riuniti in modo relativamente semplice. Conseguenza di questa complementarità è che tutte le informazioni contenute nella doppia elica possono essere duplicate a partire da entrambi i filamenti. I due tipi di paia di basi formano un numero differente di legami idrogeno: A e T ne formano due, G e C tre. Per tale motivo, la stabilità del legame GC è decisamente maggiore di quello AT. Di conseguenza, la stabilità complessiva di una molecola di DNA è direttamente correlata alla frequenza di GC presenti nella molecola stessa, nonché alla lunghezza dell'elica: una molecola di DNA è dunque tanto più stabile quanto più contiene GC ed è lunga. SENSO E ANTISENSO Una sequenza di DNA è definita senso se la sua sequenza è la stessa del relativo mRNA. La sequenza posta sul filamento opposto è invece detta antisenso. Dal momento che le RNA polimerasi lavorano producendo una copia complementare, il filamento necessario per la trascrizione è l'antisenso. Sia nei procarioti che negli eucarioti vengono prodotte numerose molecole di RNA antisenso a partire dalle sequenze senso. La funzione di questi RNA non codificanti non è stata ancora completamente chiarita. Si ritiene che gli RNA antisenso possano giocare un ruolo nella regolazione dell'espressione genica. DANNI AL DNA Il DNA può essere alterato dall'azione di numerosi agenti, genericamente definiti mutageni. Tra di essi figurano ad esempio agenti ossidanti, agenti alchilani ed anche radiazioni ad alta energia.Il tipo di danno causato al DNA dipende dal tipo di agente: gli UV, ad esempio, danneggiano il DNA generando la formazione di dimeri di timina, costituiti da ponti aberranti che si instaurano tra basi pirimidiniche adiacenti. Agenti ossidanti come i radicali liberi o il perossido di idrogeno, invece, producono danni di tipo più eterogeneo, come modificazioni di basi (in particolare di guanine) o rotture del DNA a doppio filamento Molti agenti devono il loro potere mutageno alla capacità di intercalarsi tra due basi azotate consecutive. Gli intercalanti sono tipicamente molecole planari e aromatiche, come l'etidio.Perché un intercalante possa trovare posto tra le due basi, occorre che la doppia elica si apra e perda la sua conformazione standard. Tali modifiche strutturali inibiscono sia la trascrizione che la replicazione del DNA ed aumentano la possibilità di insorgenza di mutazioni. Per tale motivo, gli intercalanti sono considerati molecole cancerogene. In ogni caso, proprio grazie alla loro capacità di inibire trascrizione e replicazione, tali molecole sono anche utilizzate in chemioterapia per inibire le cellule neoplastiche a rapida crescita. DISPOSIZIONE DEL DNA L'avvolgimento del DNA (in rosa) attorno agli istoni (in blu) Negli eucarioti, il DNA è solitamente presente all'interno di cromosomi lineari (circolari nei procarioti). La somma di tutti i cromosomi di una cellula ne costituisce il genoma; il genoma umano conta circa 3 miliardi di paia di basi contenute in 46 cromosomi. La disposizione finale a cromosomi segue precise regole gerarchiche di impacchettamento. Nelle cellule, infatti, il doppio filamento di DNA non può essere disposto a casaccio, ma deve seguire precise regole di ordinamento. Tali accorgimenti si rivelano necessari perché la lunghezza dei filamenti di DNA è solitamente molto elevata e creerebbe seri problemi alla cellula ospite. Ad esempio, il cromosoma di Escherichia coli, il procariote più studiato nella storia della biomedicina, misura circa 1 mm. In una cellula lunga solo 2 μm, come quella di E.coli, la disposizione casuale di un cromosoma del genere potrebbe generare problemi. Se una molecola di questa lunghezza si disponesse casualmente, infatti, ci sarebbe bisogno di una cellula grande almeno 1000 volte tanto. Le modalità di impacchettamento sono differenti tra gli organismi procarioti e quelli eucarioti. STRUTTURA DEL GENOMA Nel genoma, l'informazione è conservata in sequenze di DNA chiamate geni. Negli organismi eucarioti, il DNA genomico è localizzato all'interno del nucleo cellulare, nonché in piccole quantità all'interno di mitocondri e cloroplasti. Nei procarioti, il DNA è invece racchiuso in un organello irregolare, privo di membrana, contenuto nel citoplasma, chiamato nucleoide. In molte specie, solo una piccola frazione della sequenza totale di un genoma può essere trascritta e tradotta. Alcune sequenze non codificanti ricoprono un ruolo strutturale per i cromosomi. Le regioni telomeriche e centromeriche contengono solitamente pochissimi geni, ma sono necessarie per la funzione e la stabilità dei cromosomi. Nell'uomo, grandi quantità di DNA non codificante si ritrovano negli pseudogeni, copie di geni rese inattive dalla presenza di una mutazione. Queste sequenze sono considerate come fossili molecolari, anche se esistono evidenze secondo le quali si può ipotizzare che siano una sorta di materiale grezzo necessario per la creazione di nuovi geni attraverso i processi di duplicazione genica e di evoluzione divergente. TRASCRIZIONE E TRADUZIONE Un gene è una sequenza di DNA che contiene le informazioni in grado di influire sulle caratteristiche del fenotipo dell'organismo. All'interno di un gene, la sequenza di basi di DNA è utilizzata come stampo per la sintesi di una molecola di RNA che, nella maggior parte dei casi, è tradotta in una molecola peptidica. Il meccanismo attraverso il quale la sequenza nucleotidica di un gene è copiata in un filamento di RNA è detto trascrizione ed avviene per mezzo dell'enzima RNA polimerasi. Il processo di traduzione avviene nel citoplasma, dove gli mRNA si associano ai ribosomi, ed è mediato dal codice genetico. Il ribosoma permette la lettura sequenziale dei codoni del mRNA, favorendone il riconoscimento e l'interazione con specifici tRNA, molecole che trasportano gli amminoacidi corrispondenti ad ogni singolo codone. CODICE GENETICO Il codice genetico consiste in parole di tre lettere chiamate codoni, costituite dalla sequenza di tre nucleotidi (ad esempio ACT, CAG, TTT), ognuna delle quali è associata ad un particolare amminoacido. Ad esempio la timina ripetuta in una serie di tre (TTT) codifica per la fenilalanina. Utilizzando gruppi di tre lettere si possono avere fino a 64 combinazioni diverse (43), in grado di codificare per i venti diversi amminoacidi esistenti. Poiché esistono 64 triplette possibili e solo 20 amminoacidi, il codice genetico è detto ridondante (o degenere). REPLICAZIONE La divisione cellulare, necessaria ad un organismo per crescere, richiede una duplicazione del DNA cellulare, in modo che le cellule figlie possano avere la stessa informazione genetica della cellula madre. La struttura a doppia elica del DNA permette un meccanismo estremamente semplice per la replicazione del DNA. I due filamenti, infatti, sono separati e da ognuno viene creato un filamento complementare, ad opera di un enzima chiamato DNA polimerasi. Per iniziare la replicazione, occorre anzitutto l'apertura della forca replicativa, attraverso la parziale denaturazione del DNA a doppia elica, portata a termine dalle elicasi:sono enzimi che separano attivamente i due filamenti usando l'energia dell'ATP; Nelle molecole di DNA circolari dei procarioti si ha una sola regione di origine della replicazione dalla quale partono due forche replicative (la struttura prende il nome di bolla di replicazione). Quando le due forche si incontrano dal lato opposto la replicazione è completata. Negli eucarioti la replicazione di ogni cromosoma inizia invece in più punti. Le DNA polimerasi, enzimi capaci di costruire una nuova catena solo in direzione 5'-3', sono stati individuati per la prima volta da Arthur Kornberg. Dal momento che le DNA polimerasi sono in grado di svolgere la loro attività solo in direzione 5'-3', esse hanno messo a punto diversi meccanismi per copiare i due filamenti della doppia elica. Un filamento (chiamato filamento veloce) può essere replicato in modo quasi continuo, man mano che viene esposto, l'altro (filamento lento) risulta invece disseminato da brevi filamenti di DNA di nuova sintesi (i frammenti di Okazaki), ognuno dei quali presenta un innesco iniziale di RNA. I nuovi filamenti devono essere quindi completati mediante la rimozione degli inneschi da parte di endonucleasi e il riempimento degli spazi rimasti ad opera di polimerasi di riparazione. Successivamente tutti questi frammenti di DNA di nuova sintesi del filamento lento vengono legati dalle DNA ligasi. PROTEINE CHE LEGANO IL DNA Le proteine strutturali che legano il DNA sono esempi delle interazioni aspecifiche tra DNA e proteine. All'interno dei cromosomi, il DNA è associato a complessi di natura proteica, che si organizzano tra loro a formare una struttura compatta chiamata cromatina. Negli eucarioti, questa struttura presuppone il legame del DNA a piccoli complessi proteici basici chiamati istoni. Nei procarioti, invece, sono coinvolti diversI tipi di differenti proteine. Gli istoni formano un complesso a forma di disco chiamato nucleosoma. ENZIMI CHE MODIFICANO IL DNA Nucleasi e ligasi Le nucleasi sono enzimi in grado di tagliare filamenti di DNA, dal momento che catalizzano l'idrolisi del legame fosfodiesterico. Le DNA ligasi sono enzimi in grado di riunire filamenti di DNA precedentemente tagliati o spezzati, utilizzando energia chimica proveniente da ATP o da NAD. Topoisomerasi ed elicasi Le topoisomerasi sono enzimi che presentano sia un'attività nucleasica che una ligasica. Queste proteine sono in grado di modificare le proprietà topologiche del DNA. Le elicasi sono proteine in grado di utilizzare l'energia chimica presente nei nucleosidi trifosfato, soprattutto ATP, per rompere i legami idrogeno che si instaurano tra le basi azotate, permettendo l'apertura della doppia elica di DNA in singoli filamenti Polimerasi Le polimerasi sono enzimi che sintetizzano catene polinucleotidiche a partire dal nucleosidi trifosfato.Nel sito attivo di questi enzimi, il nucleoside trifosfato si appaia ad un nucleotide presente su un filamento usato come stampo: ciò permette alle polimerasi di sintetizzare in modo accurato filamenti fedelmente complementari agli stampi. Le polimerasi sono classificate sulla base del tipo di stampo che utilizzano.
Scarica