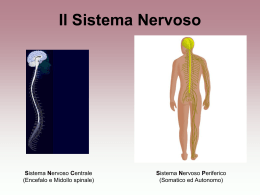
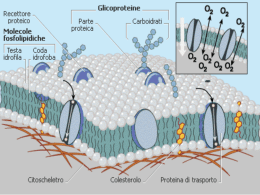
”Una porzione di tessuto cerebrale è una rete intricata che reagisce ai segnali elettrici e chimici nel tempo e nello spazio tridimensionale; essa emette e riceve configurazioni dinamiche di segnali e a tali configurazioni reagisce: queste si influenzano a vicenda e, mediante altre connessioni nervose, influenzano l’azione di altri organi del corpo, il cuore, i reni, i polmoni, i muscoli e le ghiandole. Il cervello è un controllore centrale e i suoi schemi ritmi modificano la respirazione, l’attività cardiaca, la digestione e il movimento”. Edelman, premio Nobel Per sistema nervoso si intende un'unità morfo-funzionale caratterizzata da un tessuto altamente specializzato nell'elaborazione e nella propagazione di segnali bioelettrici. Un'altra caratteristica fondamentale del sistema nervoso sta nella sua duplice collocazione anatomica: distinguiamo un nevrasse, allocato all'interno di un rivestimento costituito di tre strati fibrovascolari (meningi) situato all'interno del cranio e del canale vertebrale, e un sistema extranevrassiale che comprende tutte le strutture che decorrono all'esterno dello scheletro osseo Il sistema nervoso può essere suddiviso in due sezioni principali: sistema nervoso cerebro-spinale e sistema nervoso autonomo. Il primo è costituito dal sistema nervoso centrale e dal sistema nervoso periferico. Macroscopicamente distinguiamo i seguenti organi del SISTEMA NERVOSO CEREBRO-SPINALE Centrale il cervello il cervelletto il tronco dell'encefalo il midollo spinale encefalo Periferico le radici spinali i plessi e i gangli prevertebrali e paraverebrali i tronchi nervosi o nervi propriamente detti, con i loro rami terminali e collaterali. Le parti in cui si trovano i corpi cellulari, i dendriti ed il tratto iniziale degli assoni non rivestito da mielina costituiscono la cosiddetta sostanza grigia. Questa occupa la parte periferica degli emisferi cerebrali e del cervelletto e la regione centrale del midollo spinale. Le parti del sistema nervoso centrale che contengono le fibre nervose costituiscono la sostanza bianca. Questa si trova nella parte centrale degli emisferi cerebrali e del cervelletto e la porzione periferica del midollo spinale. SISTEMA NERVOSO AUTONOMO Il sistema nervoso autonomo controlla la muscolatura viscerale (muscolatura liscia e muscolatura cardiaca) e la secrezione ghiandolare. Inoltre è formato da due componenti: sistema nervoso simpatico (toraco-lombare) e parasimpatico (cranio-sacrale). Nel sistema nervoso periferico e in quello autonomo i corpi cellulari tendono a raccogliersi insieme formando i gangli. COMPONENTI CELLULARI DEL TESSUTO NERVOSO Il tessuto nervoso è composto essenzialmente da due tipi di cellule perenni: i neuroni, veri responsabili della ricezione e della trasmissione degli impulsi nervosi, e le cellule della glia, o nevroglia, che hanno funzioni di supporto strutturale e funzionale rispetto ai neuroni. Vengono poi talvolta impropriamente considerate come parte del tessuto nervoso alcune cellule connettivali di natura fibrosa, che offrono un sostegno strutturale al tessuto nervoso. • Neuroni - Unità funzionale del tessuto nervoso – Corpo del neurone – Dendriti - ricevono segnali elettrici e chimici da altri neuroni – Assoni - trasmettono segnali elettrici e chimici ad altri neuroni • Cellule gliali – Più abbondanti dei neuroni • Schwann, oligodendrociti, microglia, cellule ependimali, astrociti – Astrociti • Materia grigia: filtro di materiale (barriera ematoencefalica) • Materia bianca: sostegno (eccessivi in sclerosi multipla) COMPOSIZIONE DEL TESSUTO NERVOSO Il tessuto nervoso presenta un elevato contenuto di acqua che varia tra le diverse aree del sistema nervoso centrale (cervello: sostanza grigia 85%, sostanza bianca 70%) La maggior parte delle parti solide è costituita invece da lipidi: fosfolipidi sfingomieline galattolipidi (cerebrosidi e solfatidi) colesterolo Il tessuto nervoso è, dopo quello adiposo, il tessuto a più alta concentrazione lipidica Colesterolo Cerebroside Un altro importante componente del tessuto sono le proteine, che mediano diverse funzioni: funzioni strutturali attraversano la membrana cellulare costituendo i canali coinvolti nel controllo del flusso ionico della cellula riconoscono altre superfici cellulari o sostanze chimiche (recettori) sono dotate di attività enzimatica La maggior parte delle proteine della guaina mielinica sono costituite da una frazione proteolipidica e da una proteina basica FORMA E DIMENSIONE Le cellule nervose hanno forma molto variabile ma risultano costituite sempre da un corpo cellulare, il SOMA, che contiene i vari organuli e il nucleo; per tale ragione il soma viene anche detto PIRENOFORO. Dal corpo cellulare si dipartono poi i DENDRITI e l’ASSONE (o neurite), che formano rispettivamente il polo recettivo e il polo trasmittente del neurone. I dendriti possono essere considerati come una estensione del soma; essi sono in numero variabile da 3-4 fino a 20 circa. Il neurite si stacca dal soma in un punto particolare detto cono di emergenza. Si suddivide distalmente in rami molto sottili e termina con piccole dilatazioni bottoniformi soma dendr iti asson e STRUTTURA Il neurone è delimitato da una membrana cellulare che avvolge il soma e i prolungamenti. Il pirenoforo è costituito da una massa di citoplasma, denominato neuroplasma o pericarion, che circonda il nucleo e accoglie vari organuli. Nella cellula nervosa si osservano due formazioni caratteristiche: • La sostanza cromofila o zolle di Nissl che corrispondono al reticolo endoplasmatico rugoso tipico delle cellule nervose • Le neurofibrille che compongono il citoscheletro del neurone. Sono aggregati di un gran numero di sottili filamenti, i neurofilamenti, del diametro di circa 8-10nm. Accanto ai neurofilamenti, si osservano microtubuli che sono orientati con il loro terminale (+) rivolto verso la periferia e con quello (-) rivolto verso il pirenoforo. Esistono anche i microfilamenti, polimeri di actina simile ma non identica a quella presente nella cellula muscolare con un diametro di 5-6nm. INVOLUCRI DEI NEURONI: Tutte le parti delle cellule nervose sono avvolte da cellule di rivestimento rappresentate da cellule gliali quali cellule di Schwann e cellule satelliti nel SNP, oligodendrociti e astrociti nel SNC. Esse hanno il compito meccanico di proteggere e sostenere il neurone, partecipando inoltre agli scambi metabolici ed elettrolitici essenziali per il trofismo, la generazione e la propagazione degli impulsi nervosi. L’assone è circondato dalla guaina mielinica che rappresenta un buon isolante elettrico in grado di accelerare la conduzione degli impulsi nervosi. Questo perché la mielina riveste in maniera discontinua gli assoni e risulta formata da una successione di segmenti internodali, compresi tra due strozzature dette nodi di Ranvier. La mielina risulta costituita da lipidi (70%) e proteine(30%). L’alto contenuto in lipidi rappresenta una barriera per l’acqua e gli ioni idrosolubili, favorendo la funzione di isolamento della mielina. FUNZIONI DELLE CELLULE GLIALI • Produzione della guaina mielinica; • Rapida captazione e quindi all’ inattivazione dei neuro trasmettitori rilasciati dai neuroni; • Formazione del tessuto cicatriziale dopo una lesione; • Rimozione di frammenti cellulari per morte o lesione di cellule nervose (fagocitosi); • Controllo della composizione del fluido extracellulare (livelli K+); • Funzione nutritiva per le cellule nervose; • Sostegno passivo dei neuroni. Processo bidirezionale di trasferimento attivo di materiale citoplasmatico da e verso il pirenoforo (3 volte il contenuto pirenoforo/die). Avviene su un sistema di microcanalicoli costituiti dall’assemblaggio parallelo di neurofilamenti e microtubuli; essi sono associati soprattuto all’assolemma. Il citoplasma del neurite, assoplasma, contiene tubuli di SER, mitocondri, neurofilamenti e neurotubuli, tutti orientati longitudinalmente. Le sostanze per il ricambio provengono dal pirenoforo; si è visto che molte scorrono alla velocità di 1-6 mm al giorno e costituiscono il flusso assonico lento. Ci sono poi altre sostanze, genericamente definite “mediatori chimici” (come le catecolamine, ad esempio la noradrenalina), contenute in vescicole che vanno anche 100 volte più veloci (in entrambi i sensi), che costituiscono il flusso assonico rapido; provengono dal Golgi o da invaginazioni dell’estremità distale dell’assolemma e vengono trasportate dai neurotubuli. I neurotubuli hanno la funzione specifica di trasporto delle vescicole mediante proteine motrici delle loro membrane. La chinesina trasporta vescicole cariche di mediatori chimici verso l’estremità +, distale, la dineina riporta le vescicole scariche al pirenoforo, sotto forma di corpi multivescicolari. Il neurite ha il compito di ricevere l’impulso dal pirenoforo e di convogliarlo a distanza, per trasmetterlo ad altri neuroni a livello di strutture specializzate definite sinapsi. FLUSSO ASSONICO Flusso lento: proteine strutturali ANTEROGRADO proteine citoplasmatiche e di membrana precursori di neurotubuli e neurofilamenti organuli citoplasmatici. Flusso veloce: proteine funzionali trasmettitori chimici ed enzimi in vescicole Flusso veloce RETROGRADO recupero di cataboliti e di sostanze endocitate dalla terminazione assonica diffusione virus (herpes simplex, virus della rabbia) La CHINESINA e la DINEINA sono MOTORI ATP-asici citoplasmatici associati ai MICROTUBULI. La cinesina è responsabile del flusso anterogrado (dall’estremità – all’estremità + del microtubulo) la dineina di quello retrogrado (da + a -). carico adattatore chinesine dineine Gli impulsi nervosi sono trasmessi da cellula a cellula attraverso le sinapsi che permettono il collegamento di ogni neurone con altri neuroni o con cellule di tipo diverso come il muscolo scheletrico, muscolo liscio o ghiandole endocrine. Il termine di trasmissione sinaptica, pertanto, comprende la serie di eventi che costituiscono nel sistema nervoso la base della comunicazione intercellulare. I complessi meccanismi che controllano o modulano questi eventi, sia a breve che a lungo termine sono profondamente coinvolti in tutti gli aspetti della funzione cerebrale, dalle primitive funzioni autonome al comportamento conscio, all’apprendimento e alla memoria. MODULAZIONE DELLA TRASMISSIONE SINAPTICA Nelle strutture sinaptiche esistono numerosi sistemi fosforilanti, le chinasi, che sono regolati attraverso il meccanismo della fosforilazione reversibile delle proteine sinaptiche. La fosforilazione modifica le proprietà funzionali delle proteine enzimatiche e non. Le principali proteine chinasi meglio caratterizzate nelle strutture sono le seguenti: CaM=calmodulina CHINASI LOCALIZZAZIONI SINAPTICHE STRUTTURA Proteina chinasi Citosol e membrana A pre - e postsinaptica AMPcdipendente Tetrametro:2 subunità catalitiche 2 subunità regolatrici Proteina chinasi Citosol presinaptico I Ca 2+/CaMdipendente Dimero:2 subunità maggiori Proteina chinasi Citosol presinaptico II e membrana Ca2+/CaMpostsinaptica dipendente Dodecamero : 2 subunità maggiori e una subunità minore, in proporzioni diverse Famiglia delle proteine chinasi C Monomero :con variazioni della sequenza del peptide. Citosol presinaptico e membrane Nella sintesi e nel rilascio dei neurotrasmettitori La fosforilazione che modula la trasmissione sinaptica è a diversi livelli Nelle funzioni recettoriali Nell’apertura dei canali ionici Nella modulazione a lungo termine della trasmissione Le sinapsi si distinguono in : SINAPSI CHIMICHE: liberazione di neurotrasmettitori che inducono depolarizzazione della membrana plasmatica. SINAPSI ELETTRICHE: poco frequenti nei mammiferi, sono realizzate mediante giunzioni comunicanti che permettono il libero flusso di ioni; come è noto le giunzioni gap servono a mediare l’accoppiamento elettrico tra cellule adiacenti. La trasmissione dell’impulso è più veloce delle sinapsi chimiche. Le sinapsi elettriche sono presenti in particolare nelle strutture della corteccia cerebrale e nell’ippocampo. E’ “rigida”, non modificabile, non richiede la mediazione dei neurotrasmettitori chimici, in quanto le correnti generate da un ipulso nella terminazione presinaptica passano direttamente nel neurone successivo attraverso i ponti protoplasmatici che costituiscono le giunzioni comunicanti. Nelle sinapsi elettriche lo spazio che separa le cellule presinaptiche da quelle postsinaptiche è molto ristretto, di soli 2 nm. L’apprendimento e la memoria non avrebbero potuto svilupparsi in un sistema nervoso che avesse avuto soltanto sinapsi elettriche. La sinapsi chimica è di natura plastica e quindi la sua attività e la sua efficacia possono essere modificate con modalità diverse: a livello presinaptico, modulando i livelli di neurotrasmettitore liberato, a livello postsinaptico regolando l’attività dei recettori e dei canali ionici. Si identificano attraverso diverse caratteristiche peculiari tra cui la presenza di un numero notevole di vesciche raggruppate nelle terminazioni assonali presinaptiche che contengono i neurotrasmettitori o mediatori chimici. CONDUZIONE DELL’IMPULSO NERVOSO Gli impulsi che decorrono lungo le fibre nervose sono di natura elettrica esono l’espressione di una particolare attività della membrana plasmatica del neurone. SENSITIVE o AFFERENTI: conducono al nevrasse le informazioni riguardanti l’organismo o il mondo esterno FIBRE NERVOSE MOTRICI o EFFERENTI: elaborano queste informazioni e conducono agli organi effettrici gli impulsi per regolarne l’attività. LA CAPACITA’ DI CONSENTIRE LA TRASMISSIONE DEGLI IMPULSI SOLTANTO IN UNA DETERMINATA DIREZIONE E’ UNA PRIORITA FONDAMENTALE DELLA SINAPSI Tutte le cellule viventi nervose e non, hanno un piccolo eccesso di molecole cariche negativamente all’interno rispetto all’esterno che genera una differenza di potenziale fra i due lati della membrana cellulare che varia da -40 a -70 mV da cellula a cellula. Questa differenza di potenziale è definita POTENZIALE DI RIPOSO o POTENZIALE DI MEMBRANA. Questo potenziale è generato dalla presenza di anioni proteici non diffusibili all’interno della cellula e da un meccanismo di trasporto attivo(pompe ioniche) che mantiene una maggior concentrazione di ioni potassio e una minor concentrazione di ioni sodio e di ioni cloro nell’interno del neurone rispetto all’ambiente esterno. . Quando un neurone viene eccitato la membrana si depolarizza; se il voltaggio raggiunge un certo valore soglia può aprire canali per il Na+ (voltaggio dipendenti) e innescare una transitoria ulteriore caduta del potenziale che viaggia lungo l’assone con il nome di POTENZIALE D’AZIONE. La caduta del potenziale elettrico è tale da portare un inversione del potenziale di riposo (la superficie interna della membrana diventa cioè positiva e quella esterna negativa). Dopo un periodo di circa 100 microsecondi la membrana ritorna spontaneamente impermeabile al sodio per chiusura dei canali, ma aumenta la permeabilità al K+ e questo ione fuoriesce dalla cellula ripolarizzando la fibra nervosa. Il lavoro delle pompe sodio/potassio ripristina poi più lentamente la situazione iniziale. PROPAGAZIONE DEL POTENZIALE DI AZIONE Canali ionici “gated” per il passaggio di ioni secondo gradiente, specifici dei tessuti eccitabili CANALI IONICI VOLTAGGIO-DIPENDENTI •Membrana polarizzata: Canali chiusi Na+ e K+ non passano •Membrana depolarizzata: Canali aperti Na+ e K+ passano seguendo il gradiente di concentrazione (t <1 msec) •Inattivazione del canale con membrana depolarizzata: Canali inattivi Na+ e K+ non passano. Quando inattivi, non possono riaprirsi (periodo refrattario) La velocità di conduzione del potenziale d’azione dipende dal calibro della fibra. Nelle fibre mieliniche il potenziale d’azione “salta” da un nodo di Ranvier al successivo perché i canali voltaggio – dipendenti per il Na+ sono esclusivamente concentrati ai nodi (Teoria Saltatoria) FUNZIONE DELLE SINAPSI Chiarite la natura dell’impulso e le modalità con cui esso è condotto lungo l’assone occorre discutere la trasmissione dell’impulso dal neurone presinaptico a quello postsinaptico. Come già detto questa trasmissione nelle sinapsi elettriche è diretta; nelle sinapsi chimiche (la quasi totalità) richiede invece la mediazione dei neurotrasmettitori. IL TRASFERIMENTO DEL SEGNALE DALLA STRUTTURA PRESINAPTICA A QUELLA POSTSINAPTICA AVVIENE ATTRAVERSO L’ESOCITOSI DEL NEUROTRASMETTITORE CONTENUTO NELLE VESCICOLE SINAPTICHE L’EVENTO SECRETORIO E’ CALCIO-DIPENDENTE Nei bottoni terminali degli assoni sono presenti due gruppi di vescicolo soinaptiche contenenti neurotrasmettitori: 1 Quelle “agganciate” alla membrana plasmatica pronte per essere esocitate; 2 Quelle di riserva stipate nella zona attiva prossima alla membrana plasmatica. Tutte sono tenute assieme da un reticolo formato da una fosfoproteina fibrosa, la SINAPSINA con finzioni citoscheletriche. La depolarizzazione della membrana dovuta all’arrivo del potenziale di azione apre in quella sede canali per gli ioni calcio( potenziale dipendenti) che liberano ioni calcio nel bottone sinaptico. Gli ioni calcio: • inducono la fosforilazione della sinapsina così le vescicole si sganciano dal reticolo fibroso; • Si legano alla SINAPTOGAMINA (proteina presente sulla membrana della vescicola sinaptica) inducendo l’esocitosi della vescicola stessa. GLI IONI CALCIO: 1. Inducono la fosforilazione della sinapsina così le vescicole si sganciano dal reticolo fibroso; 2. Si legano alla SINAPTOGAMINA(proteina presente sulla membrana della vescicola sinaptica) inducendo l’esocitosi della vescicola stessa. LA MEMBRANA DELLE VESCICOLE SINAPTICHE VIENE RECUPERATA E RICICLATA In assenza di un meccanismo di recupero delle vescicole si dovrebbe infatti verificare, se il nervo è stimolato a lungo, un enorme aumento della membrana presinaptica. La spiegazione di questi fenomeni è fornita dall’esistenza di una micropinocitosi compensatoria nella membrana presinaptica. Le vescicole riciclate si ricaricano di neurotrasmettitori e vengono riutilizzate;quelle invecchiate sono trasportate per via retrograda verso il corpo dove sono distrutte e sostituite. I recettori per i neurotrasmettitori appartengono a due classi: 1 RECETTORI CHE SONO ESSI STESSI DEI CANALI IONICI A CONTROLLO DI LIGANDO (IONOTROPICI) che si aprono, per cambiamento conformazionale , a seguito dell’interazione con il neurotrasmettitore, permettendo il flusso di ioni secondo gradiente. Questo porta a un rapido cambiamento di potenziale della membrana postsinaptica (eccitamento). 2 RECETTORI ACCOPPIATI A PROTEINA G (RECETTORI METABOTROPICI) presenti in sinapsi che regolano funzioni che non richiedono risposte immediate ma invece prolungate nel tempo. Il loro funzionamento richiede una sequenza di eventi di trasduzione del segnale. La proteina G, attivata, può direttamente legarsi a canali ionici presenti nella membrana ed aprirli o attivare la formazione di secondi messaggeri che a loro volta influenzano la conduttanza di canali ionici. L’apertura di questi è dunque indotta indirettamente. • UNO STESSO NEUROTRAMETTITORE, LEGANDOSI A RECETTORI DIVERSI, PUO’ EVOCARE RISPOSTE RAPIDE O LENTE • IL NEUROTRAMETTITORE UNA VOLTA RILASCIATO, VIENE RAPIDAMENTE RIMOSSO O IDROLIZZATO • LE SINAPSI POSSONO ESSERE ECCITATORIE O INIBITORIE IN FUNZIONE DI QUALE CANALE IONICO VIENE APERTO. Il neurotrasmettitore delle sinapsi inibitorie, legandosi al recettore postsinaptico, provoca l’apertura di canali ionici favorenti il flusso di ioni cloro dall’esterno all’interno del neurone o ioni potassio in direzione opposta che contrasta l’attività depolarizzante espletata dalle sinapsi eccitatorie sullo stesso neurone. I PIU’ COMUNI NEUROTRASMETTITORI ECCITATORI SONO L’ACETILCOLINA E IL GLUTAMMATO, MENTRE I PRINCIPALI NEUROTRAMETTITORI INIBITORI SONO IL GABA E LA GLICINA. TRASMISSIONE DELL’IMPULSO NERVOSO Gli impulsi nervosi vengono trasmessi da cellula a cellula attraverso le sinapsi, le quali permettono il collegamento di ogni neurone con altre cellule uguali o diverse come il muscolo scheletrico, il muscolo liscio o una ghiandola endocrina. La trasmissione sinaptica comprende pertanto la serie d’eventi che costituiscono la base della comunicazione intercellulare del sistema nervoso modulata dalla fosforilazione reversibile delle proteine sinaptiche ad opera delle chinasi. La trasmissione sinaptica viene modulata dalla fosforilazione a diversi livelli: - nella sintesi e nel rilascio di neurotrasmettitori; - nell’apertura dei canali ionici; - nella modulazione a lungo termine della trasmissione. TIPI DI SINAPSI: - ELETTRICHE: rigide, senza neurotrasmettitori chimici, le correnti passano attraverso i ponti citoplasmatici delle giunzioni comunicanti; - CHIMICHE: plastiche, la loro attività e la loro efficacia possono essere modificate sia a livello presinaptico agendo sul neurotrasmettitore liberato, sia a livello postsinaptico regolando l’attività dei recettori dei canali ionici. Si identificano attraverso la presenza di vescicole nelle terminazioni neuronali che si fondono con la membrana della cellula conseguente. E’ un meccanismo che dipende dall’aumento delle concentrazioni di calcio nelle terminazioni sinaptiche ad opera della sinaptotagmina, una proteina calciofissatrice presente sulla membrana delle vescicole presinaptiche. Il segnale trasmesso dalle sinapsi chimiche parte sempre da un primo segnale elettrico che si trasforma poi in chimico per ridiventare elettrico. Il principio dell’impulso elettrico sta nel cosiddetto potenziale di riposo di membrana (-40/-70 mV), costituito da una maggiore concentrazione di ioni sodio e cloro all’esterno della membrana, e ioni potassio con ioni organici negativi all’interno della membrana. Se uno stimolo esterno viene rivolto alla superficie della membrana di una cellula nervosa fino ad abbassarne il potenziale ad un livello critico si verifica un’aumentata permeabilità ionica della membrana , dovuta ad una modificazione conformazionale delle proteine dei canali degli ioni sodio con conseguente apertura degli stessi ed un passaggio di ioni sodio all’interno e ioni potassio all’esterno. Questa modifica provoca un’inversione di potenziale sulla superficie interna della membrana fino ad un valore di 40mV con il rilascio di un neurotrasmettitore,dopodiché la permeabilità della membrana torna ad un valore normale attraverso un meccanismo che richiede energia (pompa Na/K ATPasi). CALCIO-CALMODULINA • Nella maggior parte dei processi in cui interviene il calcio lo fa legato alla calmodulina. • La colmodulina è formata da una sola catena polipeptidica con quattro siti di legame per Ca++. • LA CALMODULINA NON HA ALCUNA INTRINSECA ATTIVITA’ ENZIMATICA, MA E’ IN GRADO DI REGOLARE L’ATTIVITA’ DI MOLTI ENZIMI: FUNGE DA MEDIATORE. • Il calcio può essere considerato un secondo messaggero nelle relazioni pre e post sinaptiche. Esso può accelerare la fosforilazione di proteine cellulari attraverso l’azione dele chinasi Ca++ dipendenti e può regolare il metabolismo di AMPc. • I neurotrasmettitori che utilizzano il calcio come secondo messaggero, idrolizzano i fosfatidilinositoli di membrana. NEUROTRASMETTITORI Sono le molecole che veicolano le informazioni fra le cellule del sistema nervoso, essendo i protagonisti delle sinapsi chimiche. I neurotrasmettitori possono essere classificati secondo diversi criteri. Possono in particolare modo essere distinte due grandi famiglie: - Neurotrasmettitori a basso peso molecolare e a rapida azione utilizzati nella maggior parte delle sinapsi chimiche - Neuropeptidi ad alto peso molecolare e ad azione più lenta mediano modifiche a lungo termine INTERAZIONE POSTSINAPTICA Ogni neurotrasmettitore rilasciato nello spazio sinaptico si può legare a un eventuale recettore presente sulla membrana postsinaptica e creare così un potenziale postsinaptico. Il POTENZIALE POSTSINAPTICO PUO’ ESSERE ECCITATORIO (depolarizzante) o INIBITORIO (iperpolarizzante). Un recettore può interagire anche con molecole simili (agonisti) a quelle per esso specifiche. Come anche una stessa molecola può avere effetti diversi a seconda della cellula a cui appartiene il recettore; ad esempio l’acetilcolina produce un effetto eccitatorio nel muscolo scheletrico e un effetto inibitorio nel muscolo cardiaco. Esisitono due classi di recettori: IONOTROPICI e METABOTROPICI RECETTORI IONOTROPICI Sono canali ionici regolabili costituiti da proteine in grado di modulare il flusso ionico attraverso la membrana, e quindi il potenziale d'azione. Il legame col neurotrasmettitore causa la variazione della conformazione del canale con conseguente depolarizzazione o iperpolarizzazione. Si trovano nel sistema nervoso centrale e producono una risposta veloce, responsabile della trasmissione sinaptica eccitatoria rapida. Agiscono inoltre come sensori di segnali fisiologici. RECETTORI METABOTROPICI Il recettore metabotropico, una volta legato il ligando, avvia una serie di reazioni a cascata intracellulari mediata da un secondo messaggero ed alla base della trasduzione del segnale. Si trovano in molte parti del sistema nervoso autonomo periferico e in alcune zone del SNC, garantendo per lo più una risposta lenta e duratura. Esistono essenzialmente due tipi di risposte a questi recettori • Neurotrasmettitore → Recettore → G-protein →Adenilato ciclasi → cAMP → PKA attiva • Neurotrasmettitore → Recettore → G-protein → Fosfolipasi C → IP3/DAG → mobilitazione di calcio e attivazione di PKC ACETILCOLINA (Ach) E' rilasciata da sinapsi dette colinergiche. E’ coinvolta nelle funzioni: •Di tutte le terminazioni nervose pregangliari; •Di tutte le terminazioni nervose postgangliari del sistema parasimpatico; •Delle giunzioni neuromuscolari; •Della parte midollare della ghiandola surrenale; •Delle terminazioni nervose simpatiche postgangliari delle ghiandole sudoripare Interagisce con due tipi di recettori: muscarinici (prevalentemente nelle terminazioni parasimpatiche postgangliari) e nicotinici (nelle sinapsi gangliari del sistema simpatico e parasimpatico). SINTESI Ach E' catalizzata dall'enzima acetilcolina transferasi (CAT) e consiste nella condensazione di un acetilCoA con una molecola di colina dipendendo quindi dalla loro disponibilità. CH3CO~ S-CoA + OH-CH2-CH2-N-(CH3)3 CH3-CO-CH2-CH2-N-(CH3)3 acetil CoA colina acetilcolina + CoASH CoASH MECCANISMI DI REGOLAZIONE Le forme solubili del CAT si legano reversibilmente alle membrane dei sinaptosomi. Dopo la sintesi l’ACh viene immagazzinata in vescicole di deposito unitamente ad ATP e vesciculina (proteoglicano). Viene rilasciata quando un potenziale d’azione raggiunge le terminazioni nervose. Le vescicole di ACh si fondono con la membrana delle terminazioni e rilasciano il proprio contenuto. L'acetilcolina viene poi degradata da acetilcolinesterasi, liberando acetato e colina. Quest'ultima viene poi riassorbita dal neurone presinaptico. CATECOLAMINE noradrenalina (NA) o norepinefrina e adrenalina o epinefrina Neurotrasmettitori presenti nei nervi simpatici postgangliari e del SNC. Sono secreti dalle ghiandole surrenali. La NA è immagazzinata nelle terminazioni nervose come: •Frazione insolubile o Complesso granulareÈ costituito da NA legata ad ATP e proteine (cromogranine). Inoltre sono presenti ioni metallici (Mg2+, Ca2+ e Cu2+). •Frazione libera citoplasmatica facilmente rilasciabile in seguito ad un impulso o alla somministrazione di farmaci. SINTESI Monossido di azoto L’ossido di azoto (NO) è chiamato fattore rilassante dell’endotelio per il suo ruolo nel rilassamento della muscolatura liscia vascolare. Grazie a questo l’NO può regolare il flusso sanguigno cerebrale a seconda dei livelli di attività metabolica dei neuroni. Si forma: -nel cervello; -nell’innervazione neuronale dell’ipofisi posteriore; -nelle fibre autonome della retina; -nei corpi cellulari e nelle fibre nervose del plesso mesenterico dell’intestino; -nella midollare del surrene -nelle cellule endoteliali vascolari; -nelle cellule neuronali (in questo caso funziona come un neurotrasmettitore) La biosintesi endogena dell’NO avviene per conversione enzimatica dell’azoto guanidico della 2arginina, che in presenza di ossigeno molecolare si trasforma in citrullina; questa reazione è catalizzata dalla NO sintasi (NOS) e sono necessari NADPH, FMN e FAD. Esistono due tipi di NOS: -la forma costitutiva Ca++/CaM-dipendente induce una bassa e transiente produzione di NO da parte dell’endotelio (per modulare il tono vascolare, regolare la pressione sanguigna e l’aggregazione piastrinica); -la forma inducibile Ca++/CaM-indipendente catalizza una più lenta ma più abbondante sintesi di NO. Neuronale Inducibile Endoteliale 58 L’NO è un gas con emivita molto breve (5-6 secondi) che diffonde rapidamente attraverso le membrane cellulari ed è una molecola molto reattiva. Forme affini dell’NO hanno effetti neuroprotettivi e neurotossici. ossido nitrico (NO) L’NO può esistere come catione nitrossido (NO+) anione nitrossilico (NO-) L’NO reagisce rapidamente con l’ossigeno molecolare e con il radicale anione superossido formando l’anione perossinitrito che si decompone rapidamente in diossido di azoto e nel radicale idrossilico, uno dei più potenti ossidanti che reagisce con ogni componente cellulare. 60 61 62 Serotonina •È un’ammina biogena presente nel sistema nervoso centrale a piccole concentrazioni (prodotta dalla ghiandola pineale). •Nel circolo sanguigno in parte è legata ad albumina, in parte è libera. •Numerose tipologie di recettori situati nel cuore, nella muscolatura liscia, nel cervello. •La degradazione (nella fessura sinaptica) è ad opera della monoaminoossidasi e della n-acetil transferasi. Una bassa concentrazione di serotonina è associata all'insorgenza di depressione. L'esposizione alla luce solare e al calore del contatto umano, l'esercizio fisico, e il sorridere sono antidepressivi a costo zero che innalzano naturalmente i livelli di serotonina in circolo generando senso di benessere. La serotonina è sintetizzata partendo da L-triptofano attraverso un'idrossilazione e una decarbossilazione. Acido g-aminobutirrico (GABA) •È il principale neurotrasmettitore inibitore. •Localizzato nel: SNP, plesso mesenterico, surrene, cuore, pancreas, rene. Sintetizzato a partire da acido glutammico dalla glutammico decarbossilasi (GAD). La reazione è irreversibile e richiede PLP come coenzima ed è regolata per retroazione dal GABA. Precursori alternativi sono: arginina, ornitina e putrescina. Può essere riciclato o catabolizzato a succinato che entra nel Krebs. Il GABA è presente in scarse concentrazioni nei soggetti epilettici. Il GABA viene immagazzinato nei sinaptosomi e rilasciato in seguito a depolarizzazione recettori: GABA A: fanno parte del compleso oligomerico, che forma una canale anionico, deputato alla conduttanza di ioni cloro GABA B: rilascio neurotrasmettitori e inibizione cAMP Il GABA viene facilmente degradato in succinato che può quindi entrare nel ciclo di Krebs venendo così riciclato. Glicina E’ uno dei maggiori neurotrasmettitori inibitori del SNC ed in particolare del midollo spinale del bulbo dei mammiferi. E’ importante inoltre per il controllo sia delle funzioni motorie sia di quelle sensoriali del midollo spinale. Si trova in forma libera in tutti i tessuti dell’organismo E’ un costituente importante della dieta, ma si può anche ottenere dalla degradazione di peptidi, di proteine, di nucleotidi e di acidi nucleici. Si può formare dai carboidrati attraverso la via della 3-fosferina che porta alla formazione di serina e poi di glicina; questi due amminoacidi sono interconvertibili per azione dell’enzima serina idrossimetil transferasi In seguito al suo rilascio, la glicina si lega ai propri recettori postsinaptici, questi sono composti da due subunità una , che lega il ligando e una strutturale mentre una terza proteina, la gefirina, ha il compito di legare i recettori ai microtubuli Glutammato E’ uno dei principali neurotrasmettitori eccitatori, svolge un ruolo importante nella plasticità neuronale e nella neurotossicità; è inoltre coinvolto in molte funzioni neuronali che comprendono l’acquisizione della memoria, dell’apprendimento ed alcuni processi neurodegenerativi come l’epilessia e l’ictus Il glutammato è l’amminoacido più abbondante contenuto nel SNC e si forma dall’-chetoglutarato attraverso la via di transaminazione. Esso è anche sintetizzato attraverso altre vie: 1) dalla gluttamina per azioni della glutamminasi 2) dall’ornitina per azione dell’ornitina aminotransferasi attraverso la via della semialdeide glutammica 3) dalla prolina per azione della prolina ossidasi Il glutammato insieme all’aspartato si trovano nel sistema nervoso centrale e sono distribuiti in maniera ineguale fra cervello e midollo spinale. La maggior parte dei recettori del glutammato del sistema nervoso centrale dei mammiferi sono ionotropici (GluRi) mentre gli altri sono metabotropici (GluRm). L'eccesso anomalo di glutammato determina tossicità → Corea di Huntington Peptidi Oppiodi I principali peptidi oppioidi nel nostro organismo sono: le encefaline, le endorfine e le dinorfine. Queste molecole sono in grado di legarsi a recettori specifici accoppiati a proteine G che determinano un effetto inibitorio. Sono stati individuati tre tipi di recettori oppioidi: μ, k e δ. μ: genera analgesia (a livello sovraspinale), diminuzione attività gastro-intestinale, euforia; depressione respiratoria, k: genera analgesia (a livello spinale), depressione respiratoria, disforia; δ: non genera analgesia, ma diminuisce il transito intestinale e deprime il sistema immunitario. La sintesi dei peptidi oppioidi ha origine da una degradazione enzimatica di proteine con un peso molecolare più elevato, generalmente inattive. I tre precursori endogeni geneticamente distinti che producono peptidi multipli con distinte attività biologiche sono: - la proopiomelanocortina (POMC) - la proencefalina A - la proencefalina B (Prodinorfina) Peptidi Oppiodi Sono recettori altamente specifici per la morfina e per altri oppiodi regolatori della percezione del dolore, della temperatura, dell’appetito e dei vari stati emozionali. Si conoscono almeno sei gruppi: 1) peptidi formati da precursori dell’encefalina, costituiti da 5 amminoacidi come la Met-encefalina e la Leu-encefalina 2) peptidi formati da precursori dell’encefalina, ad esempio quelli che originano dalla proencefalina della midollare delle surrenali (Met-encefalina-Arg6-Fenil7, peptide E, dinorfina ed e -neoendorfina) 3) -endorfina ed altre encefalite correlate, le e le g endorfine 4) peptidi del tipo -casomorfina presente nel latte di mucca 5) Kiotorfina, che si ritiene sia legata alla stimolazione della biosintesi dell’encefalina e ne impedisce la degradazione 6) Demorfina Sono stati identificati tre precursori degli oppioidi che originano da tre geni distinti. Esso hanno origine attraverso una degradazione enzimatica di proteine con un peso molecolare più elevato, generalmente inattive e sintetizzate sotto il controllo dell’RNAm a livello ribosomiale. I tre precursori endogeni geneticamente distinti che producono peptidi multipli con distinte attività biologiche sono: - la proopiomelanocortina (POMC) - la proencefalina A - la proencefalina B (Prodinorfina) La POMC è un grande peptide sintetizzato dall’ipofisi e nell’ipotalamo da cui derivano ATCH, la b e g lipotropina, la b endorfina, l’MSH e il peptide corticotropina-simile del lobo intermedio (CLIP).La proenecefalina A è sintetizzata in vari siti del sistema nervso, nell’ipofisi e nella midollare delle surrenali. Contiene oltre a 4 coppie di Met-encefalina e una coppia di Leuencefalina, anche una coppia di metaencefalina con tre amminoacidi aggiuntivi.La proencefalina B, isolata dall’ipofisi anteriore, dalle ghiandole surrenali e dall’ipotalamo, contiene tre coppie di Leu-encefalina e sequenze di peptidi endogeni più grandi, la dinorfina, la neoendorfina e la rinorfina. Nonostante l’eterogeneità dei diversi tipi di recettori gli oppioidi sono coinvolti direttamente nell’inibizione del rilascio di neurotrasmettitori attraverso meccanismi che riducono l’influsso di Ca++ nelle terminazioni nervose ed inoltre inibiscono l’adenilico ciclasi e aumentano la conduttanza degli ioni K+ Metabolismo glucidico ed energetico Il sistema nervoso richiede un elevato e continuo metabolismo energetico. Il cervello estrae circa il 50% dell’ossigeno e il 10% del glucosio dal sangue arterioso. L’ossigeno passa dal sangue al cervello per diffusione, mentre il glucosio attraversa la BBB ad opera di un trasportatore. 73 La dipendenza del cervello dal glucosio si evidenzia in modo drammatico nell’ipoglicemia, quando la caduta del livello ematico del glucosio porta ad una minore captazione di ossigeno con comparsa di uno stato confusionale e poi del coma. Un substrato metabolico alternativo al glucosio ematico è il glucosio endogeno immagazzinato come glicogeno, ma il suo contenuto totale nel tessuto cerebrale è assai piccolo, e in assenza di glucosio esogeno può sostenere l’attività metabolica solo per pochi minuti. Li glicogeno è immagazzinato soprattutto negli astrociti. In condizioni di aumentata richiesta energetica ve è una rapida mobilizzazione dei depositi di glicogeno cerebrale. La glicogenolisi è stimolata da neurotrasmettitori (noradrenalina, 5idrossitriptamina, ecc.) e da altri agenti, come il potassio e l’adenosina, che vengono rilasciati nello spazio extracellulare durante l’attività neuronale. 74 La reazione limitante il catabolismo del glucosio è la sua fosforilazione, reazione irreversibile, catalizzata da esochinasi e inibita dal suo prodotto, il glucosio-6-P. Quest’ultimo substrato può essere ossidato nel ciclo dei pentosi oppure convertito in frutosio-6-P e successivamente metabolizzato attraverso la glicolisi e il ciclo degli acidi tricarbosilici. Nel tessuto nervoso, oltre il 90% del glucosio è indirizzato verso la glicolisi. 75 L’esochinasi ha un alta affinità per il glucosio e agisce rapidamente anche a basse concentrazioni di glucosio. L’esochinasi si trova in forma libera nel citosol oppure in forma legata alle membrane mitocondriali. Quando la disponibilità dei substrati è sufficiente a soddisfare la domanda metabolica, ve è un equilibrio fra l’enzima libero e quello legato. L’enzima legato ai mitocondri è più attivo, in quando è meno sensibile all’inibizione da parte di glucosio-6-P. L’enzima viene anche inibita dall’ADP, e in modo allosterico dall’acido 3-foafoglicerico, da vari nucleotidi fosfati e dall’AMPc. Il grado di legame dell’enzima alle membrane mitocondriali è inversamente correlato al rapporto ATP/ADP, cosicché quando l’utilizzazione di energia supera la disponibilità, l’equilibrio si sposta verso la forma legata, determinando quindi un’attivazione della via glicolitica. Questo meccanismo consente all’ATP di fungere da un lato come substrato dell’enzima, e dall’altro come un regolatore che riduce la produzione di ATP, esercitando la sua insufficienza sull’enzima legato. 76 Tutti gli enzimi della via glicolitica funzionano non solo nel corpo cellulare del neurone, ma anche nell’assoplasma, in particolare nelle terminazioni assonali. La glicolisi può essere aerobia od anaerobia. Il tessuto cerebrale, anche quando è ben ossigenato, produce una piccola quantità di acido lattico, compresa fra il 5-15% del glucosio totale utilizzato, che viene rimosso dal sangue venoso. L’accumulo di acido lattico nel sistema nervoso centrale è maggiore che in altri organi, come il cuore o il muscolo scheletrico, in quando l’acido lattico ha una limitata capacità di attraversare la BBB. Il metabolismo cerebrale mostra un’eterogeneità regionale, per cui, l’utilizzazione del glucosio da parte della sostanza grigia è circa tre volte maggiore di quella della sostanza bianca. 77 Il cervello non interrompe mai la sua enorme richiesta di energia. Anche durante il sonno non si ha alcuna riduzione dell’attività metabolica cerebrale, anzi questa aumenta notevolmente nella fase REM (Rapid Eyes Movements). 78 Una parte del glucosio captato, circa l’8-10%, è metabolizzato attraverso il ciclo dei pentosi, indispensabile per la produzione dei NADPH necessari alla sintesi dei lipidi. Il flusso dei pentosi fosfati diminuisce nei periodi di maggiore richiesta di energia e aumenta quando è maggiore la richiesta di lipidi, come ad esempio durante la mielinizzazione. Nel tessuto nervoso il glucosio viene anche utilizzato nella biosintesi di altre importanti molecole: il glucosio rappresenta la principale fonte di atomi di carbonio per i lipidi ed è il precursore di amminoacidi e di alcuni neurotrasmettitori (Ach, GABA, acido glutammico). 79 Il sistema nervoso centrale non viene considerato un tessuto insulino-dipendente, ma l’ormone è largamente distribuito a concentrazioni da 10 a 100 volte maggiori di quelle del plasma. Poiché il passaggio dell’insulina attraverso la BBB è molto lento e solo parziale, è stata avanzata l’ipotesi che l’insulina cerebrale possa originarsi anche nel cervello stesso. In diverse regioni del cervello sono stati identificati specifici recettori dell’insulina, che pero sono strutturalmente differenti rispetto a quelli dei tessuti bersaglio periferici. Il ruolo dell’insulina nel cervello non è ancora ben definito. 80 I lipidi nel tessuto nervoso 81 Composizione lipidica del tessuto nervoso (1) Quello nervoso è uno dei tessuti più ricchi di lipidi, essi rappresentano circa il 57% del peso secco totale della sostanza bianca e circa il 30% della sostanza grigia. Si ritiene che la composizione lipidica cerebrale costituisca un compartimento tendenzialmente stabile sia quantitativamente che qualitativamente, data la sua estrema importanza fisiologica, scarsamente influenzabile da fattori esterni. 82 Composizione lipidica del tessuto nervoso (2) I lipidi del tessuto nervoso rivestono una notevole importanza come componenti fondamentali della struttura cellulare e tissutale. 83 Composizione lipidica del tessuto nervoso (3) Inoltre, alcuni lipidi di membrana, come i fosfoinositidi, hanno un ruolo importante nella trasduzione del segnale a livello delle membrane cellulari (IP3 e DAG). 84 Composizione dei lipidi del tessuto nervoso dell’uomo Mielina (%) Sostanza bianca (%) Sostanza grigia (%) Colesterolo 27,7 27,5 22,0 Galattolipidi 27,5 26,4 7,3 Cerebrosidi 22,7 19,8 5,4 Solfatidi 3,8 5,4 1,7 Fosfogliceridi 43,1 45,9 69,5 Fosfatidiletanolamina 15,6 14,9 22,7 Fosfatidilcolina 11,2 12,8 26,7 Fosfatidilserina 4,8 7,9 8,7 Fosfatidilinositolo 0,6 0,9 2,7 Plasmalogeni 12,3 11,2 8,8 Sfingomielina 7,9 7,7 6,9 85 Il colesterolo (1) E’ l’unico sterolo presente nel cervello adulto presente in quantità significative; quello esterificato con acidi grassi si presenta in concentrazioni molto basse. Il sistema nervoso biosintetizza colesterolo a partire da AcetilCoA seguendo una via metabolica comune ad altri tessuti, tale biosintesi è particolarmente attiva durante la fase della mielinizzazione. 86 Il colesterolo (2) E’ stato dimostrato che durante lo sviluppo del cervello, i corpi chetonici, in particolare l’acido β-idrossibutirrico, sono specifici precursori del colesterolo. 87 I cerebrosidi Per cerebroside si intende un composto con una struttura in cui al gruppo alcolico – CH2OH della ceramide si lega un monosaccaride che può essere il galattosio o il glucosio. Tutti i cerebrosidi del cervello adulto sono galattocerebrosidi. 88 I solfatidi Sono galattocerebrosidi a cui è legato un addizionale gruppo solfato al carbonio 3 dell’unità glicidica: esso è definito come ceramide-galattoso-SO4¯. 89 I fosfogliceridi (1) I fosfolipidi sono i componenti più abbondanti presenti nelle cellule neuronali e la loro biosintesi avviene utilizzando come substrato il glicerolo secondo lo schema riportato. 90 Sintesi dell’acido fosfatidico CH2OH ESOSO GLICOLISI CHOH NADH NAD+ CH2-OP α GLICEROLO-P CH2OH C=O ADP ATP DH CH2-OP DHAP CH2OH CINASI CHOH CH2OH GLICEROLO 2 ACIL-CoA ACIL-TRANSFERASI 2HS-CoA O O CH2O-C-R R’-C-O-CH CH2O-P ACIDO FOSFATIDICO 91 I fosfogliceridi (2) L’acido fosfatidico, il più semplice fosfogliceride, è un intermedio centrale nelle vie di biosintesi dei fosfogliceridi e dei lipidi neutri. La sua concentrazione è bassa nel tessuto nervoso, ma, ha un alto turnover metabolico. 92 I fosfogliceridi (3) I principali fosfogliceridi derivano dal fosfatidato mediante la formazione di un legame estere tra il gruppo fosforico del fosfatidato e il gruppo ossidrilico di uno tra i numerosi alcoli che possono formare queste molecole. Nei lipidi cerebrali gli alcoli sono: etanolamina, colina, serina ed inositolo. 93 94 95 I fosfogliceridi (4) Nella sintesi di fosfatidilcolina e fosfatidiletanilamina, l’acido fosfatidico, dopo essere defosforilato da una fosfatasi, reagisce direttamente con la CDP-colina o con la CDP-etanolamina. 96 97 I fosfogliceridi (5) La fosfatidilserina ed il fosfatidilinositolo si sintetizzano a partire da CTP ed acido fosfastidico con formazione di CDP-digliceride, che, in presenza di serina o inositolo, si trasforma nel fosfolipide corrispondente. 98 Le sfingomieline (1) La sfingomielina è un fosfolipide presente nelle membrane e non deriva dal glicerolo. Lo scheletro della sfingomielina è la sfingosina, un amminoalcol contenente una lunga catena idrocarburica insatura. 99 Le sfingomieline (2) Nella sfingomielina il gruppo amminico dello scheletro di sfingosina è legato ad un acido grasso da un legame amidico. Il gruppo ossidrilico primario della sfingosina è esterificato con la fosforilcolina. 100 101 Metabolismo lipidico nell’encefalo (1) Nel tessuto nervoso sono praticamente assenti trigliceridi e acidi grassi. Gli acidi grassi si trovano nel tessuto nervoso soprattutto come componenti dei lipidi complessi. Nel tessuto nervoso vi sono anche piccole ma significative quantità di acidi grassi liberi non esterificati. Essi hanno un particolare interesse per il loro rapido turnover ed il loro metabolismo dinamico. 102 Metabolismo lipidico nell’encefalo (2) Sebbene il tessuto nervoso abbia la capacità di sintetizzare gli acidi grassi in situ, esso assorbe ed utilizza anche acidi grassi del circolo ematico, e ciò è particolarmente necessario per quanto riguarda gli acidi grassi essenziali (acido linoleico e linolenico). 103 Metabolismo lipidico nell’encefalo (3) Gli acidi grassi in circolo non rappresentano, tuttavia, una riserva di energia per il cervello perché sono legati all’albumina del plasma e quindi non sono in grado di passare la barriera ematoencefalica. In condizioni di carenza alimentare i corpi chetonici prodotti dal fegato sostituiscono parzialmente il glucosio come fonte di energia. 104 La chetogenesi (1) I corpi chetonici si formano a partire da AcetilCoA quando predomina la degradazione degli acidi grassi (in condizione di carenza glucidica). 105 La chetogenesi (2) L’Acetil-CoA entra nel ciclo dell’acido citrico soltanto se la degradazione dei grassi e quella dei carboidrati sono bilanciate. Infatti l’entrata dell’Acetil-CoA nel ciclo dipende dalla disponibilità di OAA per la formazione di citrato, ma la concentrazione di OAA è abbassata se i carboidrati non sono disponibili. 106 La chetogenesi (3) Nello stato di digiuno o nello stato diabetico l’OAA viene consumato per formare glucosio, quindi non è disponibile per la condensazione con l’Acetil-CoA; in queste condizioni l’AcetilCoA viene deviato verso la formazione di acetoacetato e di D-3-idrossibutirrato che, assieme all’acetone, sono detti corpi chetonici. 107 Metabolismo dei corpi chetonici(1) Nel cervello i corpi chetonici sono un importante combustibile. Il cervello si adatta all’utilizzo di acetoacetato durante il digiuno e nello stato diabetico. Durante il digiuno protratto il 75% del fabbisogno di combustibile del cervello è soddisfatto dai corpi chetonici. 108 Metabolismo dei corpi chetonici (2) L’acetoacetato è attivato ad Acetil-CoA mediante il trasferimento del gruppo CoA dal succinilCoA. La scissione ad opera delle tiolasi produce due molecole di Acetil-CoA che entrano nel ciclo dell’acido citrico. 109 Metabolismo dei corpi chetonici (3) In sostanza i corpi chetonici sono equivalenti energetici degli acidi grassi che riescono ad attraversare facilmente la barriera ematoencefalica. In questa fase il cervello richiede 40 g di glucosio al giorno in confronto ai 120 g richiesti nel primo giorno di digiuno. La trasformazione degli acidi grassi in corpi chetonici da parte del fegato ed il loro uso efficiente da parte del cervello diminuisce quindi le richieste di glucosio. 110 Metabolismo dei corpi chetonici (4) CH3 CH3 C=O C=O CH2 S-CoA ACETIL-CoA COO¯ ACETOACETATO Tiolasi CoA transferasi Succinil-CoA Succinato CH3 C=O CoA CH2 C=O S-CoA ACETOACETIL-CoA 111 METABOLISMO AMMINOACIDI NEL TESSUTO NERVOSO 112 La composizione del pool degli amminoacidi liberi nel cervello non è correlata ai livelli degli amminoacidi liberi presenti nel plasma o nel liquido cefalorachidiano ed è dominato dai derivati dell’acido glutammico e aspartico. Poiché i singoli amminoacidi possono variare la loro concentrazione nel cervello, nella misura del 50% o più, si ritiene che ciò possa essere riconducibile allo stretto rapporto fra l’acido glutammico e l’intensità del metabolismo glucidico, e in particolare all’attività del Ciclo di Krebs. 113 114 La maggior parte degli aminoacidi presenti nel cervello è fornita dal sangue, ma dato che le cellule nervose sono poco permeabili a molecole cariche e idrosolubili, come gli amminoacidi, sono presenti dei meccanismi specifici di trasporto attivo. 115 Particolari proteine si riscontrano nella guaina mielinica: queste includono una proteina basica e una inusuale proteina proteolipidica, solubile nei solventi organici, numerose glucoproteine lipidiche evidentemente funzionali alla struttura. Sappiamo che la caratteristica peculiare del neurone è quella di possedere un lungo assone che si estende dal corpo cellulare. È quest’ultimo l’unica sede dove sono presenti in alta concentrazione i ribosomi (sostanza di Nissl), il che indica un elevato tasso di sintesi proteica. Una delle proteine più interessanti in quanto determina la crescita e il percorso del nervo è il “fattore di crescita del nervo” (NGF). 116 FORMAZIONE DELL’AMMONIACA Le concentrazioni di ammoniaca nel cervello (0,1-0,3 μmol/g di peso fresco) sono strettamente correlate al suo stato funzionale; infatti l’attività cerebrale, la stimolazione elettrica e molti agenti convulsivanti aumentano sensibilmente i livelli di ammoniaca nel cervello. Se questa ammoniaca non viene allontanata per mezzo della detossificazione epatica, può passare la barriera ematoencefalica, aumentando i livelli di ammoniaca nel cervello con gravi conseguenze. 117 Le proteine, gli acidi nucleici e le esosamine sono le fonti dell’ammoniaca cerebrale. La desaminazione ossidativa delle amine primarie, il catabolismo della glicina, la desaminazione delle purine, delle pirimidine e della glucosamina6P contribuiscono a mantenere i livelli normali di ammoniaca del cervello. La formazione di ammoniaca cerebrale avviene per azione dell’adenilico deaminasi, che desamina l’adenosima monosfosfato, il cui gruppo aminico si è formato per trasferimento del gruppo NH2 dagli amminoacidi attraverso l’acido glutammico e aspartico. 118 IL CICLO DELL’UREA E L’ALLONTANAMENTO DELL’AMMONIACA Il catabolismo degli aminoacidi genera ammoniaca e ioni ammonio, che sono tossici, in particolare per il sistema nervoso centrale. La maggiorparte dell’ammoniaca è resa innocua a livello dei siti di formazione, in misura minore l’azoto entra in circolo, nella vena porta, come ammoniaca o come alanina, che sono utilizzati dal fegato per la produzione di urea. 119 Nel cervello, in particolare nelle cellule gliali l’ammoniaca viene in gran parte trasformata in glutammina, che a sua volta è rilasciata nel circolo sanguigno, consentendo di mantenere concentrazioni basse di questo composto. L’allontanamento dell’ammoniaca si rende necessario perché altera la trasmissione sinaptica, sia eccitatoria che inibitoria. 120 121 Il ciclo dell’urea rappresenta la via principale di eliminazione dell’azoto dall’organismo. Nei neonati la mancanza di uno dei qualsiasi enzimi del ciclo causa iperammoniemia, che altera il funzionamento del cervello e provoca uno stato patologico noto come ENCEFALOPATIA. Questo problema si manifesta entro le prime 48 ore di vita e inevitabilmente è aggravato dall’assunzione di un alimento ricco in proteine, come il latte. 122 Nutrienti e funzione cerebrale La disponibilità di alcuni nutrienti ha notevoli effetti sulla funzionalità cerebrale e può influire sul comportamento: - Zinco: stimola la formazione di GABA ed una sua carenza ha profondi effetti sul comportamenti; - Rame: una sua carenza influisce sul normale sviluppo del cervello; - Selenio: mantiene la capacità antiossidante del cervello; - Glucosio: precursore dei neurotrasmettitori; - Amminoacidi: una carenza di triptofano, da cui deriva la serotonina provoca un’aumentata sensibilità a stimoli dolorosi, la tirosina è indispensabile per la sintesi delle catecolamine; - Vitamine: la carenza di una qualsiasi delle vitamine idrosolubili comporta sempre dei sintomi a livello neurologico. 123
Scaricare