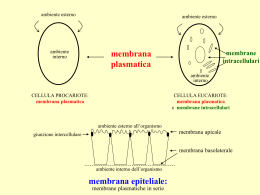
LA MEMBRANA PLASMATICA - Deve trattenere i materiali in soluzione nella cellula in modo che essi non filtrino nell’ambiente esterno - Deve consentire i necessari scambi di materiali verso l’interno e verso l’esterno della cellula - La struttura molecolare della membrane e le molteplici funzioni che esse svolgono nella vita della cellula. - Trasporto di soluti attraverso le membrane. COMPOSIZIONE CHIMICA LIPIDI: Fosfolipidi-sfingolipidi Colesterolo Glicolipidi PROTEINE: Semplici Glicoproteine Lipoproteine Enzimi ( 5’nucleosidasi; ATPasi; fosfatasi alcalina; adenilato ciclasi; fosfomonoestaerasi) CARBOIDARTI: acido sialico In acqua le interazioni delle code idrofobiche e delle teste idrofiliche generano un doppio strato fosfolipidico. Le teste sono dirette verso l’esterno, dove interagiscono con l’acqua che le circonda. Le code sono rivolte verso l’interno. Unità lipidiche a una coda Modello di Gorter e Grendel non spiega le proprietà della membrana connesse a tensione superficiale Permeabilità dei soluti Resistenza elettrica DATI IN DISACCORDO CON IL MODELLO DI DANIELLI E DAVSON Lo spessore della membrana plasmatica è di 6-8nm. Di questi, 4-5nm sono costituiti dallo strato lipidico, per cui le proteine devono essere contenute nei restanti 1-2nm Non spiega le peculiarità dei diversi tipi di membrana Le teste idrofile dei lipidi dovevano essere protette dalle proteine superficiali all’azione delle fosfolipasi La maggior parte delle proteine era insolubile in acqua, poteva essere estratta solo con l’impiego di solventi organici o detergenti esperimenti evidenziavano la natura fluida della membrana: proteine e lipidi si muovevano liberamente dentro il piano delle membrane GPI: glicosilfosfatidilinositolo Le proteine periferiche di membrana si collegano, mediante forze elettrostatiche deboli e legami idrogeno, alle superfici della membrana con le porzioni idrofile delle proteine integrali e probabilmente con le teste polari dei lipidi di membrana. Queste proteine possono essere estratte mediante variazioni del pH o della forza ionica. Le proteine ancorate ai lipidi sono unite con legame covalente a molecole lipidiche incluse nell’uno o nell’altro dei due strati della membrana. Sono legate ad un acido grasso o ad un derivato dell’isoprene chiamato gruppo prenile. Numerose proteine situate sulla superficie esterna sono connesse con legame covalente al glicosilfosfatidilinositolo, un glicolipide del monostrato esterno della membrana. Esse possono essere rilasciate dalla membrana mediante l’azione dell’enzima fosfolipasi C. Le proteine di membrana differiscono per la loro mobilità Le particelle proteiche portano una netta carica negativa SDS, sodiododecilsolfato PROTEINE DI MEMBRANA E LORO FUNZIONI Enzimi [es. glucosio fosfatasi (RE) gliceraldeide-3 fosfodeidrogenasi (GPD)] Proteine che trasportano elettroni (es. citocromi, ferroproteine) Proteine di trasporto (es. aa e proteine) Proteine canale ATP-asi di trasporto Recettori Connessoni (gap junction) tra cellule; desmotubuli (plasmodesmi) Proteine dei pori presenti sulle membrane esterne dei mitocondri e cloroplasti; i pori dell’involucro nucleare Proteine che svolgono un ruolo nella stabilizzazione e nel modellamento della membrana cellulare (es. anchirina, spettrina, proteina della banda 4.1 ) GLI ACIDI GRASSI Sono indispensabili per la strutture e la funzione della membrana poiché le loro code di idrocarburi costituiscono un’efficace barriera idrofoba nei riguardi della diffusione dei soluti polari La maggior parte degli acidi grassi delle membrane ha una lunghezza tra 12 e 20 atomi di carbonio (più frequenti quelli a 16-18 atomi di carbonio) Oltre alle differenze nella lunghezza delle code di idrocarburi, variano anche per la presenza e il numero di doppi legami • Dimostrazione della mobilità dei lipidi all’interno delle membrane. • Tecnica: RECUPERO DELLA FLUORESCENZA DOPO FOTOSBIANCAMENTO I lipidi sono distribuiti asimmetricamente tra i due monostrati interno ed esterno(es. glicosfingolipidi solo nel monostarto esterno; PE, PC, PS, sono più abbondanti nel monostrato interno) LE MEMBRANE FUNZIONANO CORRETTAMENTE SOLO ALLO STATO FLUIDO LA FLUIDITA’ DELLA MEMBRANA CAMBIA CON LA TEMPERATURA: diminuisce quando la temperatura si abbassa, aumenta quando la temperatura si innalza. TEMPERATURA DI TRANSIZIONE (Tm) = la temperatura alla quale un doppio strato lipidico gelifica “congela” se raffreddato e diventa di nuovo fluido (fonde) se riscaldato Il cambiamento dello stato della membrana è denominato TRANSIZIONE DI FASE Per funzionare correttamente una membrana deve essere mantenuta allo stato fluido, cioè a temperatura al di sopra del suo valore di Tm. La fluidità di una membrana dipende soprattutto dai tipi di lipidi che contiene(in particolare dalla lunghezza delle catene degli acidi grassi e il loro grado di insaturazione) La temperatura alla quale un acido grasso saturo fonde aumenta con l’aumentare delle lunghezza delle catene. Le membrane che contengono parecchi acidi grassi saturi hanno Tm elevate, mentre le membrane con acidi grassi principalmente insaturi hanno Tm basse. I Lipidi della maggior parte delle membrane contengono acidi grassi che variano sia per la lunghezza che per il grado di insaturazione della catena. Questo assicura che le membrane, a temperature fisiologiche, si mantengano allo stato fluido. La maggior parte degli organismi, sia procarioti che eucarioti, è capace di regolare la fluidità della membrana, principalmente variandone la composizione lipidica (adattamento omeoviscoso dei poichilotermi: batteri, funghi, protisti, piante, animali a sangue freddo). I GLICOLIPIDI e le GLICOPROTEINE di superficie sono responsabili di: Riconoscimento cellulare Adesione cellulare (nei tessuti e organi) Trasporto attraverso la membrana Interazione tra le cellule Inibizione proliferazione da contatto Proprietà delle membrane Definiscono confini esterni di cellule e organelli cellulari e regolano il traffico di molecole e ioni attraverso questi confini. Sono: •Superfici di separazione tra compartimenti •Superfici di riconoscimento come recettori per ormoni •Superfici catalitiche Struttura delle membrane •simili a fogli (6-10 nm) •costituite da lipidi e proteine •non presentano legami covalenti tra le molecole componenti •asimmetriche •fluide La composizione proteica delle membrane da fonti diverse varia più della composizione lipidica. Le membrane plasmatiche contengono molte glicoproteine, quelle degli organelli intracellulari raramente.
Scarica