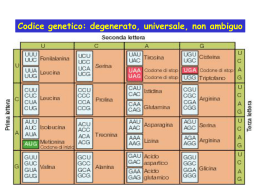
I R I BOSOM I I RIBOSOMI sono organuli citoplasmatici presenti in tutte le cellule, sia procariotiche che eucariotiche. Sono visibili al M.O. solo quando presenti in gran numero, (come capita nelle cellule nervose), ed appaiono come ammassi citoplasmatici di materiale basofilo. Nelle sezioni istologiche esaminate al M.E. si presentano come particelle dense con diametro di circa 15-30 nm e possono essere distribuiti nella matrice ialoplasmatica (RIBOSOMI LIBERI) oppure essere adesi alla superficie esterna delle membrane del RER (RIBOSOMI ASSOCIATI ALLE MEMBRANE). Possono trovarsi liberi nel citoplasma oppure adesi al R.E. In entrambi i casi si possono trovare associati in gruppi, denominati Polisomi o Poliribosomi, costituiti da un esile filamento di RNA messaggero I POLISOMI possono assumere una caratteristica forma a spirale. Le ricerche con isotopi radioattivi hanno dimostrato che le sintesi proteiche avvengono soltanto SUI RIBOSOMI ASSOCIATI NEL COMPLESSO POLISOMALE E NON SUI RIBOSOMI SINGOLI, i poliribosomi quindi rappresentano le unità attive di sintesi della cellula. Si tratta di corpiccioli di natura ribonucleoproteica che nel citoplasma di tutte le cellule presiedono ai processi di sintesi proteica. L’importanza che essi rivestono è notevole dato che le interazioni tra RNA messaggero, RNA transfer e amminoacidi, sembrano dipendere in larga misura dall’organizzazione molecolare del ribosoma. I ribosomi possono essere isolati mediante centrifugazione e la loro analisi ha dimostrato che essi presentano una considerevole uniformità di dimensioni, struttura e composizione. Sui RIBOSOMI ISOLATI è possibile valutare mediante ultracentrifugazione il coefficiente di sedimentazione che è espresso in unità Svedberg o unità S. Mediante queste tecniche si è potuto stabilire che i ribosomi dei batteri hanno un coefficiente di sedimentazione di 70S che corrisponde ad un peso molecolare di 2,6x106dalton, mentre quelli delle cellule superiori sia animali che vegetali hanno un coefficiente di sedimentazione di circa 80S con un peso molecolare di 4,5x 106 dalton. Gli esperimenti hanno inoltre dimostrato che i ribosomi sono formati da due subunità diseguali. Il ribosoma 70S è costituito da una subunità maggiore di 50S e una subunità minore di 30S; il ribosoma 80S da due subunità di 60S e di 40S. L’associazione tra le due subunità è mantenuta dalla presenza di MAGNESIO fino a concentrazioni di 1mM. Abbassando tale concentrazione il ribosoma si dissocia, elevando di nuovo la concentrazione si riforma la particella ribosomiale completa. Se la concentrazione di magnesio aumenta notevolmente due ribosomi si combinano per formare un dimero. Figura 3.45 Mediante metodi di centrifugazione su gradiente è possibile separare non soltanto ribosomi singoli ma anche poliribosomi. Questi appaiono al M.E. come aggregati di un numero variabile da tre a varie decine, di singole unità, (collegate, come già detto prima, da un esile filamento di mRNA). Tale filamento è digerito dalla ribonucleasi che dissocia il poliribosoma nei ribosomi singoli e si ritiene che sia una molecola di mRNA che porta l’informazione genetica per la sintesi proteica dal DNA ai ribosomi. È stata avanzata l’ipotesi che la molecola di mRNA sia interposta tra le due subunità del ribosoma; tale configurazione può spiegare come circa 25 nucleotidi dell’mRNA polisomale siano protetti dall’azione della ribonucleasi quando il polisoma è trattato con questo enzima. La lunghezza dell’mRNA e il numero di monomeri del polisoma possono variare in misura notevole e dipendono dalla lunghezza della catena polipeptidica che ogni molecola specifica di mRNA può codificare. Sebbene non esista una dimostrazione diretta al M.E., le indagini di biologia molecolare fanno ritenere che nella cellula i ribosomi singoli siano sempre dissociati nelle due subunità e che queste si uniscano tra loro e con la molecola di mRNA formando il polisoma soltanto durante il processo di sintesi proteica, per dissociarsi al termine di esso. L’analisi chimica dei ribosomi isolati ha dimostrato che entrambe le subunità sono composte da molecole di acido ribonucleico ribosomiale (rRNA) e da proteine in proporzioni pressoché uguali. Nei procarioti il rapporto in peso tra rRNA e proteine è di 2:1, la subunità minore 30S contiene un’unica molecola di RNA 16S alla quale sono associate circa 21 proteine diverse. La subunità maggiore 50S ha una molecola di RNA 23S ed una di 5S ad esse sono legate circa 35 proteine. I ribosomi degli eucarioti sono più complessi: la subunità minore 40S contiene una molecola di RNA 18S e la subunità maggiore 60S tre molecole di RNA: una 28S, una5,8S ed una 5S le proteine ribosomiali ammontano a 80 distribuite nelle due subunità (circa 50 nella sub maggiore e 30 in quella inferiore). La maggior parte di queste proteine è di carattere basico. Nei mitocondri e nei cloroplasti si trovano ribosomi simili a quelli procariotici. Nei lieviti i ribosomi mitocondriali (o MITORIBOSOMI), sono di 70S, nei protozoi sono di 80S, ma con due subunità entrambe di 55S. Nei mitoribosomi di 55S sono presenti solo due RNA, uno di 12 S nella subunità minore ed uno di 21 S in quella maggiore. La struttura secondaria delle molecole di rRNA è poco conosciuta, ma si ritiene che esse siano costituite da una CATENA UNICA RIPIEGATA IN ALCUNE ZONE SU SE STESSA A FORMARE TRATTI A DOPPIA ELICA CON APPAIAMENTO COMPLEMENTARE DELLE BASI. La caratteristica dell’rRNA è di avere un contenuto in guanina+citosina più elevato(60%) del contenuto in adenina+uracile (40%) e di essere relativamente stabile. Tutte le molecole di rRNA, 28S,5,8S,5S e 18S sono caratterizzate dalla presenza, in aggiunta alle 4 principali basi azotate, di basi metilate, di pseudouridina e di 2-0-metilribosio L’RNA ribosomiale rappresenta la frazione più grande di RNA nel citoplasma costituendo il 70-85% dell’RNA totale. Sia nei batteri che nelle cellule superiori, la subunità maggiore del ribosoma contiene anche una molecola di RNA di modeste dimensioni 5S costituita da 120 nucleotidi. LA SUA STRUTTURA È COSTITUITA DA UN FILAMENTO UNICO RIPIEGATO IN ALCUNI PUNTI SU SE STESSO A FORMARE TRATTI A DOPPIA ELICA. La funzione di tale struttura è ancora sconosciuta. La modalità di interazione tra RNA e proteine nelle subunità non è completamente nota. Tuttavia si ritiene che la molecola di RNA e proteine ribosomiche siano tra loro intimamente associate. Le proteine interagiscono con la molecola di RNA principalmente nelle regioni a catena singola, formando un filamento nucleoproteico che a sua volta fittamente avvolto su se stesso a formare una particella compatta. È attualmente ben stabilito che i ribosomi si originano nel nucleolo. Figura 3.47 INNESCO O INIZIO DELLA SINTESI PROTEICA La fase iniziale della biosintesi proteica è definita come la sequenza di eventi che porta alla formazione di un complesso tra un ribosoma 80S, una molecola di metionina-t-RNA(met-t-RNA) ed una molecola di mRNA. Diverse proteine non ribosomiche sono necessarie per la formazione di questo complesso di inizio ed esse sono dette Fattori di inizio(IF). Un ribosoma contiene 4 siti di legame per molecole di RNA: uno per l’mRNA e tre (sito E, sito P e sito A) per il tRNA ALLUNGAMENTO DELLA CATENA POLIPEPTIDICA Dopo che si è formato il complesso d’inizio, il met-t-RNA è sul sito peptidico (sito P) del ribosoma. Un secondo aminoacil-t-RNA corrispondente alla seconda tripletta dell’mRNA si colloca sulla subunità maggiore del ribosoma legandosi al sitoaminoacidico (sito A) . Sul ribosoma sono ora localizzate due molecole di tRNA affiancate e legate a due codon consecutivi dell’mRNA; esse sono disposte l’una in posizione prossimale rispetto all’estremità 5’dell’mRNA e l’altra in posizione distale. L’attacco dell’aminoacil-t-RNA al sito A del ribosoma richiede GTP come fornitore di energia ed è promosso dal cosiddetto “fattore di allungamento”(EF1). La metionina (e nelle tappe successive la catena polipeptidica nascente) si stacca dal suo tRNA per rottura del legame estere tra l’adenosina terminale dell’RNA di trasporto ed il gruppo carbossilico dell’aminoacido; tra questo gruppo ed il gruppo aminico del secondo aminoacil-t-RNA affiancato al codon successivo si forma il legame peptidico. L’AMINOACIL-T-RNA LEGATO AL SITO A RICEVENDO LA CATENA POLIPEPTIDICA DIVENTA PEPTIDILT-RNA. La formazione del legame peptidico è catalizzata dall’enzima peptiltransferasi che è localizzato sulla subunità 60S ed è forse parte integrante del sito P. Il peptidil-tRNA si dissocia dal sitoA, rimanendo tuttavia Il sito A è in questo ancorato al suo codon, ed il ribosoma si sposta rispetto al momento vacante. Il complesso tRNA –mRNA di un codon verso l’estremità 3’ dell’mRNA, oppure l’mRNA scorre relativamente al ribosoma in direzione opposta. Come conseguenza il met-tRNA deacilato processo di traslocazione dell’mRNA relativamente al esce dal ribosoma ed il peptidil-tRNA sempre ancorato alla ribosoma e del peptidil- sua tripletta dell’mRNA viene a collocarsi sul sito P. tRNA dal sito A al sito P è forse il risultato di un cambiamento conformazionale del ribosoma che avviene a spese dell’idrolisi del GTP e richiede l’intervento del fattore proteico EF2 o traslocasi; per ogni tappa di traslocazione si forma un complesso EF2-GTP che si lega al ribosoma. Un terzo aminoacil-tRNA complementare alla terza tripletta dell’mRNA si attacca al ribosoma collocandosi sul suo sito aminoacidico vacante. Sul ribosoma sono di nuovo situati fianco a fianco due tRNA legati a due codon contigui dell’mRNA : il peptidil-tRNA che porta la catena polipeptidica nascente ed è inserito sul sito P e l’aminoacil-tRNA che porta l’ultimo aminoacidico incorporato ed è associato al sito A. Con l’attacco del nuovo aminoacil-tRNA, la situazione è ritornata a quella di partenza, con la differenza che la catena polipeptidica si è accresciuta di un aminoacido e la traduzione dell’mRNA è progredita di un codon. Il ciclo costituito dalle precedenti fasi si ripete per ogni codon, incorporando un aminoacido per volta, finché tutto l’mRNA è tradotto. sitoP sitoA TERMINAZIONE DELLA TRADUZIONE Quando l’ultima tripletta dell’mRNA alla sua estremità 3’ è stata letta, la traduzione si arresta e la catena polipeptidica si libera dal ribosoma e dal tRNA, contemporaneamente alla dissociazione del ribosoma nelle due subunità. Un fattore di natura proteica detto Fattore di dissociazione interviene nella separazione delle due subunità. Il segnale di terminazione è dato da una opiù triplette NON SENSO (UAA, UGA, UAG) cioè non traducibili mancando il rispettivo tRNA. TERMINAZIONE DELLA TRADUZIONE Quando il peptidil-tRNA è nel sito P ed una tripletta non senso di terminazione è nel sito A, un fattore proteico di rilascio RF si lega al sito A in presenza di GTP e catalizza la reazione di terminazione. Questa reazione consiste nell’idrolisi del legame che unisce il peptide al tRNA , nel loro rilascio, nel distacco dell’mRNA dal ribosoma ed infine nell’idrolisi del GTP.
Scarica