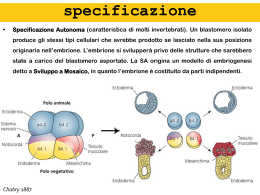
Non solo il Gradiente Morfogenetico (una molecola diffusibile agisce secondo un gradiente di concentrazione) può spiegare il differenziamento cellulare a seguito dell’esposizione alla fonte di activina. 1.Cascata di reazioni di amplificazione: l’activina agisce solo a concentrazioni elevate, inducendo il differenziamento SOLO delle cellule immediatamente adiacenti alla fonte. Tali cellule emettono un secondo segnale, che induce il differenziamento delle cellule adiacenti, e così via. Tale ipotesi è stata verificata (ed esclusa) interponendo uno strato di cellule insensibili all’activina tra l’emisfero animale (in grado di rispondere) e la fonte di activina. 2.Migrazione cellulare: l’activina induce nelle cellule immediatamente adiacenti, oltre al differenziamento, anche migrazione cellulare; le cellule differenziate si ritrovano quindi a grande distanza dalla fonte di activina dalla quale son partite. Tale ipotesi è stata verificata (ed esclusa) colorando in modo differente le cellule: Le cellule competenti per la risposta all’activina hanno circa 500 recettori activina- specifici sulla loro superficie cellulare. Se i recettori complessati dall’activina sono tra 100 e 300, viene trascritto il gene bra, e le cellule differenziano verso un destino mesodermico ventrale. Se il numero di recettori complessati è più alto, allora viene attivato gsc, che porta a differenziamento mesodermico dorsale. in senso specificazione Specificazione sinciziale (insetti). Segmentazione superficiale. Blastoderma sinciziale Gilbert, BIOLOGIA DELLO SVILUPPO specific. sinciziale Specificazione sinciziale (insetti). mRNA localizzati asimmetricamente Gradienti morfogenetici delle proteine bcd e nos Gilbert, BIOLOGIA DELLO SVILUPPO mRNA bicoid proteina bicoid Gilbert, BIOLOGIA DELLO SVILUPPO, Zanichelli editore S.p.A. Copyright © 2005 mRNA bicoid anteriore mRNA nanos posteriore mRNA hunchback e caudal distribuiti uniformemente. Gilbert, BIOLOGIA DELLO SVILUPPO, Zanichelli editore S.p.A. Copyright © 2005 (NRE) (pum) (pum) Gilbert, BIOLOGIA DELLO SVILUPPO, Zanichelli editore S.p.A. Copyright © 20 equivalenza del genoma Tutte le cellule dell ’ organismo hanno effettivamente lo stesso identico genoma? Oppure il patrimonio genetico della cellula ormai differenziata ha subito modificazioni funzionali irreversibili? delle Gilbert, BIOLOGIA DELLO SVILUPPO, Zanichelli editore S.p.A. Copyright © 2005 equivalenza del genoma Hans Spemann e la Teoria del Trasferimento Nucleare 1938: una cellula già terminalmente differenziata è in grado di riprogrammare l'informazione in termini di espressione genica e di controllare lo sviluppo embrionale? Spemann propose di prelevare il nucleo da una cellula di un embrione in avanzata fase di sviluppo e trasferirlo nel citoplasma di una cellula uovo enucleata. Spemann non pote' condurre l'esperimento per la mancanza di strumenti adatti alla manipolazione e dissezione delle cellule somatiche e germinali. equivalenza del genoma exp. Robert Briggs e Tom J. King, 1952 Rana pipiens equivalenza del genoma equivalenza del genoma Briggs e King pongono le basi sperimentali (creano quegli strumenti che erano mancati a Spemann 14 anni prima quando elaborava la teoria del Trasferimento Nucleare) per fornire le risposte ai quesiti di quegli anni: (?) Il nucleo di una cellula differenziata conserva ancora tutte le informazioni originarie; inoltre, tale nucleo risulta riprogrammabile ed in grado di originare un nuovo individuo. (?) La sinergia tra il citoplasma ospite dell’uovo e il nucleo che in esso viene rilocato è sufficiente a riattivare il nucleo a dirigere lo sviluppo del nuovo individuo. equivalenza del genoma 1962: J. Gurdon ritenta l'esperimento di Briggs e King, usando nuclei di cellule differenziate d'intestino di girino di Xeanopus laevis. 1.4 la % di successo a girino. Utilizzando il trapianto in serie - il nucleo viene trapiantato nell ’ uovo che viene lasciato sviluppare fino a blastula. Il nucleo della cellula di blastula si trasferisce in un altro uovo enucleato e così via - ottenne un successo del 7% comprendente 7 rane adulte fertili. Gurdon dimostra che i nuclei di cellule somatiche differenziate, citoplasma di modificano il genetico. un trasferiti uovo proprio nel enucleato, programma equivalenza del genoma 1997: I. Wilmut e collaboratori clonano il primo mammifero. Prelievo cellule somatiche da ghiandola mammaria di adulto (razza Finn-Dorset). Coltura cellulare e arresto in G1 (le cellule, 2n, son quindi sincronizzate). Oociti dalla donatrice (razza Scottish blackface) in seconda metafase meiotica enucleati. Fusione cellulare somatica/oocito e attivazione dell’oocito stesso mediante impulsi elettrici. Trasferimento in utero di pecora gravida. % di successo: 1/434 oociti impiantati = Dolly. Dimostrazione della totipotenza del nucleo dei mammiferi: il genoma rimane inalterato e capace di differenziare in tutti i tipi cellulari. Gilbert, BIOLOGIA DELLO SVILUPPO, Zanichelli editore S.p.A. Copyright © 2005 (TATA binding protein) Gilbert, BIOLOGIA DELLO SVILUPPO, Zanichelli editore S.p.A. Copyright © 2005 Gilbert, BIOLOGIA DELLO SVILUPPO, Zanichelli editore S.p.A. Copyright © 2005 variazioni epigenetiche Modifica ereditabile (quindi trasmissibile alla progenie), che può riguardare un cromosoma o l’attività di un gene, che non varia la sequenza nucleotidica. Es: metilazione del DNA Nei vertebrati la metilazione interessa solamente la Citosina sul dinucleotide CpG ad opera di enzimi DNA-metiltransferasi (Dnmt) che aggiungono un gruppo metile al C5 della citosina: il risultato è la 5-metilcitosina. Il 3-5% circa delle C risulta metilato in cellule eucariotiche metilazione del DNA Lo schema di metilazione viene mantenuto ad opera di DNA-metil transferasi di mantenimento, che hanno particolare affinità per le sequenze emi-metilate: tendono quindi a metilare il nuovo filamento che si è formato su uno stampo metilato. Globina embrionale e > gene attivo nell ’ embrione, inattivo nel feto Globina fetale g > gene inattivo nell’embrione, attivo nel feto Gilbert, BIOLOGIA DELLO SVILUPPO, Zanichelli editore S.p.A. Copyright © 2005 Istone acetiltransferasi (HAT) trasferiscono gruppi acetile agli istoni del nucleosoma, che viene così destabilizzato consentendo la trascrizione del gene. I fattori di trascrizione sono spesso in grado di reclutare le HAT o sono essi stessi delle HAT. Istone deacetilasi > reazione ‘inversa’ con conseguente stabilizzazione dello stato trascrizionalmente inattivo del gene. Vengono anche reclutate dalle regioni metilate. Gilbert, BIOLOGIA DELLO SVILUPPO, Zanichelli editore S.p.A. Copyright © 2005 anfibi • Facili da mantenere • Iniezione di ormone • Uova grosse e manipolabili • Sviluppo embrionale piuttosto rapido anfibi solco di segmentazione gastrulazione La gastrulazione in Xenopus ha inizio nella regione della semiluna grigia (futuro lato dorsale dell’embrione) Lo spermatozoo è importante nel determinare non il movimento in sé, quanto il senso della rotazione del citoplasma corticale. Un uovo artificialmente attivato ruota ancora il suo citoplasma, ma in direzione non prevedibile. anfibi Il citoplasma ruota grazie alla presenza di un apparato di microtubuli che si forma, al polo vegetativo, tra il citoplasma corticale e quello interno. Tale apparato compare appena prima della rotazione e scompare subito dopo. anfibi - segmentazione Uova mesolecitiche (quantità modesta di vitello al polo vegetativo), segmentazione oloblastica (totale) ineguale con simmetria radiale solco di segmentazione blastocele 1. Permette la migrazione cellulare durante i massivi movimenti della gastrulazione 2. Impedisce il contatto – quindi l’interazione – tra le cellule. emisfero (calotta) animale Nieuwkoop (1973) anfibi - segmentazione Blastocele (cavità) Morula (16-64 cellule) Blastula (128 cellule) oligonucleotidi antisenso anti-EP-caderina. Heasman et al, 1994 blastocele controllo Heasman et al, 1994 oligonucleotidi antisenso anti-EP-caderina gastrulazione gastrulazione La blastula deve ridistribuire le cellule/aree in modo tale da portare verso l’interno quelle (cellule/aree) che daranno origine alle strutture endodermiche, verso l’ esterno quelle che origineranno il rivestimento dell’ organismo e interporre a questi due “blocchi“ le cellule/aree con destino mesodermico. Løvtrup (1975), Landstrom & Løvtrup (1979), Keller (1975, 1976) anfibi localizzazione RNA materni 1. l’mRNA deve essere trasportato al polo vegetativo. 2. l’mRNA deve essere ancorato al polo vegetativo. localizzazione RNA materni Vg1LE (Vg1 localization element), una sequenza di circa 350 nucleotidi localizzata nella regione 3´UTR (untranslated region) dell’mRNA di Vg1 mRNA, necessaria affinchè il messaggero localizzi al polo vegetativo dell ’ uovo. Vg1LE è una sequenza zipcode (C.A.P.). Vg1LE viene complessata da almeno 6 proteine, 2 sole delle quali note, Vg1RBP/Vera e VgRBP60. Omologhi di Vera funzionano come localizzatori di mRNA anche in altri sistemi (es. pollo). Piccoli RNA non tradotti (Xlsirt) sono essenziali per l’associazione dell’mRNA Vg1 con la cell cortex del polo vegetativo, ma non per il trasporto. Essi contengono stretch complementari al mRNA per Vg1 e ne mediano l’associazione con la corticale. localizzazione RNA materni Vg1RBP/Vera A V Vg1RBP/Vera colocalizza con il mRNA di Vg1 ed è necessario per la sua corretta localizzazione al polo vegetativo dell’uovo polarità dell’uovo La prova sperimentale dell’imposizione di una polarità all’uovo di Xenopus da parte di determinanti citoplasmatici viene dall’ablazione dei mRNA codificanti per tali determinanti, VegT (Zhang et al, 1998) e Vg1 (Joseph & Melton, 1998). La mancanza di VegT e Vg1 altera la distribuzione delle cellule nei tre foglietti embrionali. anfibi Gilbert, BIOLOGIA DELLO SVILUPPO, Zanichelli editore S.p.A. Copyright © 2005
Scarica