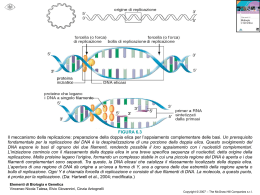
IL MITOCONDRIO FUSIONE E FISSIONE DEI MITOCONDRI I mitocondri sono organelli cellulari. Essi sono connessi con il citoscheletro e sono in uno stato dinamico nel quale costantemente si fondono e si dividono PROTEINE COINVOLTE NELLA FUSIONE E FISSIONE DEI MITOCONDRI PROTEINE DI FUSIONE PROTEINE DI FISSIONE OPA 1: Proteina dello spazio intermembrana avente attività GTPasica Fis 1: Proteina della membrana esterna Mfn: proteina della membrana esterna con un dominio GTPasico e una regione Coiled coil Drp1: Proteina citosolica e mitocondriale avente attività GTPasica TESSUTI DEI MAMMIFERI ALTERATI DA DIFETTI NELLE PROTEINE DI FUSIONE LA BIOGENESI DEL MITOCONDRIO Il mitocondrio contiene un proprio genoma e apparati distinti di replicazione, trascrizione e traduzione. Tuttavia l’mtDNA codifica solo per un ridotto numero di proteine (13) mentre il resto è codificato dal DNA nucleare LE PROTEINE CODIFICATE DALL’mtDNA SONO CONTENUTE NEI COMPLESSI RESPIRATORI IL DNA MITOCONDRIALE DEI METAZOI Costituisce circa l’ 1% del genoma: ogni cellula contiene da 103 a 104 copie di mtDNA Codifica per 2 rRNA, 13 mRNA e 22 tRNA Si trova in strutture ricche di proteine dette nucleoidi I NUCLEOIDI I nucleoidi rappresentano l’unità dinamica ed ereditaria dell’mtDNA Vertebrati : 50 - 800 nucleoidi/cellula 2 a 6 nucleoidi / organello 2 a 15 molecole di mtDNA/nucleoide PROTEINE ASSOCIATE CON I NUCLEOIDI NEI VERTEBRATI Proteine coinvolte nel mantenimento, replicazione e/o trascrizione dell’mtDNA POL G A POLG B Twinkl e MtSSB TFAM POLRMT MtTFB (Xenopus ) TFB1M (uo mo) TFB2M(uomo) Rnase MRP Copurif icazione Con mt DNA Parziale Parziale Parziale Parziale SI N.D. N.D. N.D. N.D. N.D. Distribuz ione intramitocond riale MITOCONDRI N.D. NUCLEOIDI NUCLEOIDI NUCLEOIDI N.D. N.D. MITOCONDRI MITOCONDRI N.D. Proteine con funzione sconosciuta LON proteasi BRCA1 ANT1 PDC-E2 BCKD-E2 PHB2 Copurif icazione Con mt DNA N.D. N.D. SI SI. SI SI Distribuz ione intramitocond riale N.D. NUCLEOIDI N.D. N.D. N.D. N.D. Tra le proteine contenute nel nucleoide alcune hanno un ruolo noto nel metabolismo dell’mtDNA, altre non sono apparentemente collegate all’mtDNA dato che svolgono un ruolo a se stante. RIMODELLAMENTO METABOLICO DEI NUCLEOIDI MITOCONDRIALI Studi in cellule di lievito hanno portato alla proposta di un modello di rimodellamento del nucleoide secondo il quale in seguito a cambiamenti del metabolismo cellulare (es situazioni di stress) l’aconitasi perderebbe il cluster Fe-S e si riposizionerebbe sul nucleoide, contribuendo a stabilizzare l’mtDNA MODELLO DI SEGREGAZIONE DEI NUCLEOIDI IN LIEVITO Attraverso contatti con proteine di membrana il nucleoide mitocondriale si collega al citoscheletro. In questo modo il nucleoide si muove con i mitocondri nelle cellule della progenie durante la divisione cellulare IL DNA MITOCONDRIALE DEI METAZOI •Costituisce circa l’ 1% del genoma: ogni cellula contiene da 103 a 104 copie di mtDNA •Codifica per 2 rRNA, 13 mRNA e 22 tRNA •Si trova in strutture ricche di proteine dette nucleoidi •Viene ereditato per via materna •Viene riparato con scarsa efficienza •Il processo di ricombinazione del DNA mitocondriale è poco efficiente •Nel mitocondrio esiste il fenomeno della eteroplasmia, ossia la presenza di più di un genoma mitocondriale nello stesso individuo IL BOTTLENECK La mancanza di ricombinazione del DNA mit potrebbe far pensare che il genoma mit si comporti come un organismo asessuato aploide, per cui l’accumulo di mutazioni deleterie potrebbe determinare l’estinzione dell’organismo (Muller Ratchet). Invece esistono meccanismi specifici che prevengono questa possibilità Uno di questi meccanismi è detto bottleneck o campionamento MECCANISMO DEL BOTTLENECK Il bottleneck rappresenta un esempio di replicazione rilassata del DNA mit per cui durante la citochinesi alcune molecole si replicano ed altre no. Durante i primi stadi dell’ovogenesi avviene un campionamento delle molecole di DNA mit, fino a circa 200/cellula Successivamente nell’ovocita primario il contenuto cellulare di mtDNA e di mitocondri aumenta considerevolmente fino a 100000 molecole di mtDNA/cellula. Il campionamento (bottleneck) avviene durante lo sviluppo di un feto femmina destinato a diventare la madre nelle generazioni successive. Mentre le cellule germinali primordiali non differiscono nel loro livello di eteroplasmia la drammatica segregazione dei genotipi mitocondriali che avviene durante lo sviluppo degli ovociti primari dà luogo ad un alta varianza intercellulare Il campionamento avviene in modo casuale senza meccanismi di selezione positivi o negativi CONSEGUENZE DEL BOTTLENECK Il campionamento dell’mtDNA porterebbe alla formazione di individui Omoplasmici nei quali si sarebbero perse le mutazioni debolmente deleterie prima che esse si accumulino. Ciò eviterebbe il verificarsi del “Muller ratchet” Tuttavia le mutazioni che sfuggono a questa selezione che sono debolmente deleterie e che non vengono rimosse, sarebbero amplificate e potrebbero diventare omoplasmiche Questo è probabilmente il meccanismo per la patogenesi delle malattie mitocondriali ereditate per via materna MODELLO ASINCRONO DI REPLICAZIONE DEL’mtDNA MODELLI COMPETITIVI DI REPLICAZIONE DELL’mtDNA NEI MAMMIFERI Replicazione asincrona (Clayton) Replicazione bidirezionale da siti multipli di inizio Replicazione Unidirezionale Sincrona (Holt 2000) LA TERMINAZIONE PREMATURA DELLA REPLICAZIONE DELL’ mtDNA DIPENDE DAL SUO SITO DI INIZIO L’origine localizzata in pos 57 è responsabile per il mantenimento dell’mtDNA in condizioni di steady state mentre le altre due origini sarebbero richieste per il recupero dopo la deplezione dell’mtDNA e per accellerare la replicazione dell’mtDNA in risposta a richieste fisiologiche IL DNA MITOCONDRIALE UMANO La replicazione dell’H-strand (modello Clayton) inizia nella regione non codificante e richiede la presenza di un primer ad RNA che è sintetizzato dalla RNA polimerasi mitocondriale a partire dal promotore PL. Il principale sito di inizio della replicazione si trova a livello di una sequenza conservata CSB-1 che rappresenta il punto più importante per la transizione tra l’RNA primer e l’mtDNA IL D-LOOP 5’ RNA CSBs DNA 3’ TAS Una notevole proporzione di molecole di mtDNA di mammifero contiene una struttura a tripla elica detta D-loop, costituita da un breve tratto di strand H nascente appaiata alla strand L templato, con conseguente spiazzamento della strand H materna. Le estremità 3’ del D-loop si trovano in corrispondenza di blocchi di sequenza conservate dette TAS E’ stato proposto che la struttura del D-loop rappresenti una replicazione abortiva e che il numero di copie dell’mtDNA possa anche essere regolato dall’espansione di questa struttura. FATTORI COINVOLTI NELLA REPLICAZIONE DELL’ mtDNA La replicazione dell’mtDNA dipende da numerosi fattori, ognuno dei quali svolge un ruolo importante nel processo. SINTESI DEL PRIMER Richiede: •mtRNA pol •Fattori di trascrizioneTFAM TFB1 TFB2 TFAM •Attiva la trascrizione delle strand H ed L legandosi ai rispettivi promotori HSP E LSP •Si lega al DNA anche in maniera regolare e senza specificità di sequenza svolgendo la funzione di histone-like protein •E’ essenziale per il mantenimento dell’ mtDNA in quanto la sua deplezione é letale 100 TFAM 150 200 ITH ITL 250 300 CSB3 CSB2 OH1 CSB1 OH2 350 HSP LSP 400 450 500 550 TFAM, TFB1 E TFB2 • TFB1 e TFB2 sono due fattori proteici che insieme a TFAM stimolano la sintesi del primer TFB2 TFB1 TFAM LSP mtRNA pol. • Sembra che TFB1 e TFB2 colleghino TFAM con l’RNA polimerasi facilitando l’inizio della trascrizione MATURAZIONE DEL PRIMER RICHIEDE UNA RIBONUCLEOPROTEINA (MRP) o AVVIENE IN MODO PROTEINA INDIPENDENTE L’enzima MRP è un’endonucleasi costituita da una parte proteica e da una componente ad RNA. Secondo gli autori l’enzima svolgerebbe il suo ruolo tagliando l’RNA e producendo il primer per la replicazione. Il taglio avverrebbe quando l’RNA è ibridizzato con il DNA stampo formando una struttura R-loop L’esistenza di strutture a singola strand nella regione CSB1 suggerisce che esse rappresentino dei siti di pausa della trascrizione. Pertanto la formazione del primer sarebbe indipendente dall’azione di una nucleasi Utilizzando un sistema purificato di trascrizione in vitro si è osservato che la trascrizione della strand L termina nel box CSB II. Pertanto anche questo studio sostiene che la formazione del primer è indipendente dalla presenza della endonucleasi MRP LA TERMINAZIONE PREMATURA DELLA REPLICAZIONE DELL’ mtDNA DIPENDE DAL SUO SITO DI INIZIO L’origine localizzata in pos 57 è responsabile per il mantenimento dell’mtDNA in condizioni di steady state mentre le altre due origini sarebbero richieste per il recupero dopo la deplezione dell’mtDNA e per accellerare la replicazione dell’mtDNA in risposta a richieste fisiologiche FATTORI COINVOLTI NELLA REPLICAZIONE DELL’ mtDNA mtSSB Proteina che lega il DNA a singolo filamento evitandone la rinaturazione ALTRE CARATTERISTICHE DI mtSSB • stimola l’attività 3’-5’ esonucleasica e l’attività 5’-3’ polimerasica della DNA polimerasi g. ed aumenta anche la processività dell’enzima • La deplezione di mtSSB induce la deplezione dell’mtDNA •Studi in Drosophila mostrano che l’espressione di mtSSB è regolata dal legame ad elementi DRE che controllano la replicazione del DNA di Drosophila. Ciò definisce un possibile legame tra replicazione nucleare e mitocondriale Il collegamento tra espressione di mtSSB e variazioni nel contenuto dell’mtDNA suggerisce che essa sia asssociata con meccanismi che regolano la replicazione o la stabilità dell’mtDNA ELICASI E’ stata descritta una elicasi mitocondriale 5’-3’ richiesta per la replicazionedell’ mtDNA Essa è stata denominata TWINKLE (omologa alla elicasiprimasi di T7) • L’attività di TWINKLE è stimolata da mtSSB • Mutazioni nel gene per TWINKLE sono associate con delezioni multiple nell’mtDNA di pazienti sofferenti per l’adPEO • Studi con animali transgenici e esperimenti di RNAi hanno mostrato che l’espressione di TWINKLE è strettamente associata con il livello dell’mtDNA •TWINKLE, mtSSB e TFAM sono localizzate nei nucleodi COME VIENE REGOLATA L’ESTENSIONE DEL D-LOOP ? 5’ RNA CSBs DNA 3’ TAS IPOTESI Le sequenze TAS potrebbero essere un segnale di riconoscimento per una DNA-binding protein che potrebbe funzionare bloccando la progressione della forca replicativa Questa proteina potrebbe svolgere un azione antielicasica Gli autori identificano una proteina di circa 45KDa che si lega alle regioni TAS A E TAS B del DNA mit umano mtDBP, THE D-LOOP BINDING PROTEIN IN SEA URCHIN OH P NCR Q T E 12 S F Q Cyt b ND5 ND6 mtDBP mtDBP P.lividus mtDNA mtDBP • IS A TRANSCRIPTION TERMINATION FACTOR MtDBP works as a bidirectional transcription termination factor in an in vitro heterologous transcription assay. pDBP-term(F) bmtDBP - pDBP-term(R) - RO T T mtDBP AND mtDNA REPLICATION…….. OH nascent H-strand RNA primer 12S E T mtDBP 1098 G-rich AT P Q N 1126 1175 NCR Is mtDBP involved in mtDNA replication, perhaps by impeding the synthesis of nascent DNA (“contrahelicase activity”) ? IN VITRO HETEROLOGOUS HELICASE ASSAY • SV40 large T Antigen (replicative helicase) • Recombinant purified mtDBP • Partial duplex substrates that contain mtDBP binding site alone or followed by its flanking region, in both orientations mtDBP mtDBP G-rich G-rich helicase substrates DNA-BOUND MTDBP EXHIBITS A NON-POLAR CONTRAHELICASE ACTIVITY Substrate: mtDBP binding site OH 3’ DBP.For 5’ 1090 mtDBP 100 °C - + + + + + T Ag 1246 T Ag % helicase activity 100 86 65 37 10 mtDBP OH 3’ 5’ DBP.Rev - 100 °C + + + + + T Ag 1090 1246 T Ag % helicase activity 100 95 90 44 5 MTDBP IS A DUAL FUNCTION PROTEIN: It terminates transcription bidirectionally It is a bidirectional contrahelicase that could act as a negative regulator of mtDNA synthesis. MtDBP could coordinate the interplay between replication and transcription. PROKARYOTIC FACTORS MEDIATING TERMINATION OF TRANSCRIPTION AND REPLICATION Bacterial factors Tus and RTP (D. Bastia and coll.): - block the replication fork in a polar mode (contrahelicase) - exert a polar stop to elongating RNA pol; passage of an RNA transcript from the permissive side of DNA-protein complex causes dislodging of the protein: the arrested helicase is restored. replic. replic. RNApol RNApol blocking side permissive side replic. 3’ blocking side replic. 5’ 5’ Blocked RNApol 3’ What is the mechanism that allows mtDNA synthesis to resume? MODEL Dissociation of DNA-bound mtDBP caused by the passage, in the opposite direction, of the RNA polymerase through the protein-DNA complex. IMPACT OF TRANSCRIPTION INVASION ON THE DNA-BOUND MTDBP T7 promoter mtDBP DNA template T7 RNA pol T7 RNA pol transcribes and invades mtDBP-DNA complex T7 promoter mtDBP is dislodged and trapped by a labelled DNA probe 32P mtDBP NTPs T7 RNA pol mtDBP DNA template + _ + + + + + + _ + + + + _ + + + + + + + _ _ _ + _ + _ _ mtDBP/DNA complex _ DNA:prot. 1:6 1:13 free oligonucleotide A POSSIBLE MODEL FOR THE ABROGATION OF mtDBP CONTRAHELICASE ACTIVITY arrested H-strand OH DNA helicase 12S E T G-rich AT P Q mtDBP mtRNA pol Readthrough transcription dislodges mtDBP elongating H-strand 12S E T OH G-rich AT P Q mtRNA pol Transcription invasion of the protein-DNA complex causes dislodging of mtDBP. The arrested helicase is restored and mtDNA replication can resume. DNA POLIMERASI g Nelle cellule animali la DNA polimerasi mitocondriale (DNA pol g) é una proteina costituita da due subunità: • subunità catalitica con attività esonucleasica 3’-5’ ed attività polimerasica 5’-3’ di circa 140 kDa •subunità regolatrice di 35-54 kDa La subunità regolatrice è omologa alle aminoacil-tRNA sintetasi di classe II Essa aumenta l’affinità della polimerasi per l’mtDNA e promuove un più forte legame dei nucleotidi aumentando la velocità di polimerizzazione REGOLAZIONE DEL NUMERO DI COPIE DEL DNA Il numero di copie del DNA può dipendere: da fattori direttamente coinvolti nella replicazione da fattori coinvolti nella biogenesi del mitocondrio da fattori coinvolti nel ciclo cellulare REGOLAZIONE DELLA BIOGENESI MITOCONDRIALE In risposta a danni al DNA, p53 interagisce con l’mtDNA e con la DNA polimerasi aumentando la velocità di replicazione MODELLO PER LA REGOLAZIONE DEL NUMERO DI COPIE DELL’mtDNA DA PARTE DELLA VIA DI CONTROLLO Mec1/Rad 53 DEL CICLO CELLULARE IL NUMERO DI COPIE DELL’ mtDNA DIPENDE DA FATTORI PROTEICI E DA ELEMENTI LIMITANTI COME IL POOL DEI NUCLEOTIDI MAPPA DELLE MUTAZIONI PATOLOGICHE DEL DNA MITOCONDRIALE UMANO (DI MAURO 2005) ASPETTI CLINICI, MORFOLOGICI E BIOCHIMICI DI DISORDINI COLLEGATI A MUTAZIONI DELL’mtDNA GENI NUCLEARI COINVOLTI NEL MANTENIMENTO DELL’mtDNA SOSTITUZIONI AMINOACIDICHE NELLA DNA POL g RESPONSABILI DI MUTAZIONI PATOLOGICHE O DI POLIMORFISMI Sporadic PEO Male infertility AdPEO Mutazioni autosomiche recessive SANDO Sindrome di Alper
Scarica