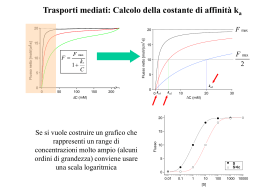
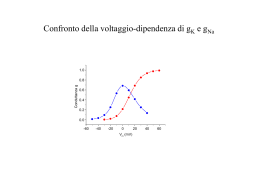
Attività elettrica dei neuroni Neuroni piramidali del SNC che stanno crescendo su una matrice di glia/fibroblasti Nel potenziale d’azione si distinguono due fasi: -fase di DEPOLARIZZAZIONE -fase di RIPOLARIZZAZIONE tempo Basi ioniche del potenziale d’azione • conduttanze e correnti di Na voltaggio-dipendenti • conduttanze e correnti di K voltaggio-dipendenti Qual è il ruolo dei canali del Na+ nella genesi del potenziale d’azione? 1. Osservazione di Hodgkin e Huxley (1939): – al picco di un potenziale d’azione il potenziale di membrana diventa transitoriamente positivo all’interno – overshoot. 2. Ipotesi: è coinvolto il Na – l’entrata di Na attraverso la membrana continuerà fino al raggiungimento del potenziale di equilibrio del Na (ENa). ENa = 58 log ([Na]o / [Na]i) = 58 log (460/50) = 59 mV 50% [Na]o -> ENa = 38 mV 33% [Na]o -> ENa = 28 nV 3. Esperimento di Hodgkin e Katz (1949): riducendo [Na]o riduzione dell’overshoot Conclusione: Il potenziale d’azione è il risultato di un grande e transitorio aumento nella permeabilità al Na della membrana. Al potenziale di riposo Al picco del PA PK : PNa : PCl 1.0: 0.04: 0.45 1.0 : 20 : 0.45 Che dire della fase di caduta del potenziale d’azione? - I canali del Na semplicemente si chiudono (inattivazione) Ma se la causa fosse solo questa, la ripolarizzazione sarebbe più lenta di quanto osservato sperimentalmente - Si verifica un successivo grosso aumento della permeabilità della membrana al K dovuto all’apertura di canali del K voltaggio-dipendenti. Il potenziale di membrana ritorna verso EK Il potenziale di membrana iperpolarizza (diventa più negativo del potenziale di riposo) per un certo tempo Per spiegare l’ampiezza e l’andamento temporale del potenziale d’azione Ipotesi: Le conduttanze del Na e K sono voltaggio-dipendenti - La probabilità che i canali si aprano aumenta con la depolarizzazione. L’effetto della conduttanza al Na - rigenerativo. Il processo -si autoalimenta -feedback positivo La depolarizzazione aumenta la conduttanza al K con un certo ritardo. Aumento di gNa Depolarizz. Corrente di Na entrante Aumento di gK -autolimitante -feedback negativo Depolarizz. Corrente di K uscente Ripolarizz. Ipotesi di Hodgkin e Katz: La depolarizzazione della cellula al di sopra del valore di soglia causa un breve aumento della permeabilità della membrana agli ioni Na. In che maniera la permeabilità di membrana (conduttanza) agli ioni Na varia con il potenziale di membrana? gNa = INa / (Vm – ENa) Esperimento: Occorrerebbe misurare l’ampiezza della corrente di Na+ (INa) che fluisce attraverso la membrana a diversi livelli del potenziale di membrana (Vm) C’è un problema: La corrente fluisce attraverso la membrana il potenziale di membrana cambia la conduttanza della membrana si modifica sia al variare del potenziale di membrana che nel tempo Cioè, esiste forte interdipendenza del PM e del gating dei canali del Na (e del K) sia il PM che la conduttanza dei canali cambiano costantemente. depolarizzazione INa entra. Per risolvere questo problema, K. Cole sviluppò la tecnica del voltage-clamp. Canali Na aprono Alcuni tipi di canali Ionici sono sempre aperti, altri no! extracell. intracell. Sempre aperti Il loro comportamento è ohmico: la relazione I/V è lineare g=costante Voltaggio-dipendenti: rispondono a variazioni di Vm Il loro comportamento non è ohmico: la relazione I/V non è lineare perché g non è costante [g=f(Vm)] Modello del gating secondo H&H Il modello di H&H della gate assume una reazione cinetica del 1o ordine tra gli stati aperto e chiuso della particella di gating a C O b Quindi, la probabilità della particella di trovarsi nello stato aperto può essere descritta da: (1-Po) a b Po Trattandosi di una cinetica del 1o ordine, sarà: dPo a (1 Po ) bPo dt Allo stato stazionario (equilibrio) sarà: a quindi, P o dPo a (1 Po ) bPo 0 dt ab Risolvendo l’equazione differenziale del 1o ordine e applicando la condizione al contorno Po0 Po(t 0), si ottiene: Po( t ) Po0 ( Po Po0 ) 1 e ( a b )t 1/(ab)=t rappresenta la costante di tempo dell’attivazione ed è un indice della velocità di attivazione della particella di gating. Se p particelle di gating indipendenti sono coinvolte nel gating del canale, allora il canale seguirà il seguente andamento temporale: Pochann (t ) Po(t )p Prob. Di Attivaz. 1.0 Po(t) 0.8 0.6 [Po(t)]p 0.4 0.2 0.0 0 2 4 Tempo 6 8 La tecnica del voltage-clamp consiste nel bloccare il potenziale di membrana ad un valore costante nel tempo e nel registrare le correnti ioniche transmembranarie generate a tale potenziale di membrana Potenziale di membrana finale (Vm) costante (potenziale di comando) (mV) 20 Potenziale di riposo (Vr) -70 0 10 20 Tempo (ms) 30 40 Corrente ionica (nA) 60 Corrente transmembranaria generata dal passaggio di Vm da –70 mV a +20 mV 40 20 0 -20 -40 -60 0 10 20 30 40 Tempo (ms) Vantaggi del voltage-clamp: In genere, nel caso di canali voltaggio-dipendenti, gm = f(V,t) ma, 1. Vm è bloccato ad un valore costante gm = f(t) soltanto e può essere dedotto dall’andamento della corrente ionica Ii 2. E’ possibile separare Ii da Ic. Infatti cm è caricata istantaneamente Voltaggio (mV) Separazione farmacologica delle correnti IK e INa 20 -70 0 10 20 Tempo (ms) 30 40 + TTX (IK) INa+IK Corrente ionica (nA) 60 40 20 0 + TEA (INa) -20 -40 -60 0 10 20 Tempo (ms) 30 40 Voltaggio-dipendenza dei canali Kv 140 120 100 + I(K ) max (nA) Voltaggio (mV) 60 50 40 30 20 10 0 -10 -20 -30 80 60 40 IK=gK• (V-EK) 20 0 -70 0 10 20 30 -20 40 -30 -20 -10 0 Tempo (ms) 10 20 30 40 50 60 Voltaggio (mV) 140 1.0 120 Conduttanza (μS) Corrente K + (nA) 100 80 60 40 0.8 0.6 0.4 gK = IK/(V-EK) 20 0.2 0 0.0 -20 0 10 20 Tempo (ms) 30 40 EK=-80 mV -30 -20 -10 0 10 20 30 40 50 60 Voltaggio (mV) Canali voltaggio-indipendenti Il potenziale di riposo di una cellula è determinato dalla presenza di due canali permeabili rispettivamente ai cationi A+ e B+ attraverso i quali passano le correnti ioniche IA e IB indicate in tabella. Dopo aver disegnato i rispettivi grafici I/V, determinare: 1) le conduttanze gA e gB; 2) se i due canali sono voltaggio-dipendenti o -indipendenti; 3) il potenziale di equilibrio dei due ioni A e B; 4) il potenziale di riposo Vr della cellula. V(mV) -120 -110 -100 -90 -80 -70 -60 -50 -40 -30 IA (nA) -1650 -1100 -550 0 550 1100 1650 2200 2750 3300 V(mV) -20 -10 0 10 20 30 40 50 60 70 IB (nA) -1320 -1100 -880 -660 -440 -220 0 220 440 660 V(mV) -120 -110 -100 -90 -80 -70 -60 -50 -40 -30 IA =gA (V-EA ) (nA) -1650 -1100 -550 0 550 1100 1650 2200 2750 3300 V(mV) -20 -10 0 10 20 30 40 50 60 70 IB=gB(V-EB) (nA) -1320 -1100 -880 -660 -440 -220 0 220 440 660 4000 3000 2000 EA 1000 0 -150 -100 -50 0 -1000 -2000 EA=-90mV EB=+40mV EB g1A=-1650/[-120-(-90)]= g1B=-1320/[-20-(+40)]= 55 mS 22 mS Vr=(E AgA+EBgB)/(g A+gB)= -52.9 mV 50 100 Quesito del giorno Un ricercatore, in seguito ad esperimenti condotti in voltageclamp ha ricavato che la corrente di K+ (IK) dipende dal potenziale di membrana (V) secondo la seguente relazione: 1 IK 4.6 (V 85) 1 (V 10) / 9 1 e (pA) 1) Calcolare i valori di IK nell’intervallo di potenziali tra –80 e +80 mV ad intervalli di 10 mV e metterli in grafico in funzione di V; 2) Dai valori di IK ricavati, e sapendo che EK= -85 mV, calcolare i valori corrispondenti di gK e metterli in grafico in funzione di V; 3) A quale valore di V sara’ aperta la meta’ dei canali? IK = 4.6·[1-1/(1+EXP((V+10/9))]·(V+85) (pA) EK = -85 mV IK 0.0 0.1 0.4 1.9 7.1 24.7 74.1 172.5 294.2 394.3 466.4 522.9 572.8 620.2 666.7 712.9 759.0 Vm -EK 5 15 25 35 45 55 65 75 85 95 105 115 125 135 145 155 165 gK 0.002 0.006 0.018 0.053 0.158 0.450 1.139 2.300 3.461 4.150 4.442 4.547 4.582 4.594 4.598 4.599 4.600 600 400 200 0 -100 -50 0 50 100 mV nS Vm -80 -70 -60 -50 -40 -30 -20 -10 0 10 20 30 40 50 60 70 80 pA 800 5.0 4.0 3.0 2.0 1.0 0.0 -100 -50 0 mV 50 100 Voltaggio-dipendenza dei canali Na 1 0 40 30 20 10 0 -10 -20 -30 -40 -50 -1 I(Na) picco (nA) Voltaggio (mV) 60 -2 -3 -4 -6 -70 -7 0 1 2 3 4 -50 -40 -30 -20 -10 0 10 20 30 40 0 -1 -2 -3 -4 -5 0.12 gNa = INa/(VENa) 0.10 ENa=+50 mV 0.14 0 1 2 Tempo (ms) 3 4 0.08 0.06 0.04 0.02 0.00 -50 -40 -30 -20 -10 0 10 20 30 40 0 1 2 Tempo (ms) 60 Voltaggio (mV) Conduttanza (μS) I(Na) (nA) I(Na) (nA) Tempo (ms) 0 -1 -2 -3 -4 -5 -6 -7 INa=gNa• (V-ENa) -5 3 4 Voltaggio (mV) Separazione della fase di attivazione della corrente di Na dall’inattivazione La pronasi rimuove l’inattivazione Voltaggio (mV) -80 -60 -40 Corrente di Na (nA) Voltaggio (mV) 40 30 20 10 0 -10 -20 -30 -40 -50 -60 -70 -80 -90 -20 0 0 20 40 -200 -400 -600 -800 0 10 20 30 40 tempo (ms) 0 2.5 -200 2.0 -400 -600 -800 -1000 1.5 0.5 1.0 0.5 -1200 0 0 10 20 tempo (ms) 30 40 0 -80 -60 -40 -20 0 Voltaggio (mV) 20 40 Prob. canale aperto Conduttanza ( m S) Corrente (nA) 1 L’ampiezza della corrente di K aumenta con la depolarizzazione. L’ampiezza della corrente di Na aumenta e poi diminuisce con la depolarizzazione. Perchè? Depolarizzazione: Depolarizzazione: IK= gK (Vm–EK) INa= gNa (Vm–ENa) La corrente attraverso il canale dipende da 1. Permeabilità del canale: conduttanza le conduttanze del Na e K sono voltaggio-dipendenti 2. Driving force – dovuta al potenziale elettrico attraverso la membrana e al gradiente di concentrazione degli ioni. Come risalire dai valori di conduttanza allo stato stazionario alla probabilità di trovare una singola gate del canale aperta Grafico della conduttanza g(V) allo stato stazionario Nel caso di un canale inattivante, esso è stato ottenuto rimuovendo l’inattivazione 2.5 Gmax 1.5 0.5 1.0 0.5 0 Prob. canale aperto C onduttanza ( m S) 1 2.0 0 -80 -60 -40 -20 0 20 g (V ) Po g P Gmax o 40 Voltaggio (mV) Ipotesi: l’attivazione del canale è regolata da x gates n uguali e indipendenti. Allora, se n∞ è la probabilità di avere la singola gate n aperta allo stato stazionario, la probabilità Po di trovare il canale aperto allo stato stazionario (in assenza di inattivazione) sarà: Po n x Probabilità composta: n·n·n…. (x volte) Quindi, dai valori di Po si può risalire ai valori di n∞: n x Po Quesito del giorno Un ricercatore, in seguito ad esperimenti condotti in voltageclamp e utilizzando la tossina di scorpione per bloccare l’inattivazione, ha ricavato che la conduttanza del canale del Na+ (gNa) dipende dal potenziale di membrana (V) secondo la seguente relazione: 1 (nS) gNa 5.6 1 (V 30) / 6 1 e 1) Calcolare i valori di gNa nell’intervallo di potenziali tra – 80 e +80 mV ad intervalli di 10 mV e metterli in grafico in funzione di V; a quale valore di gNa sarà aperta la metà dei canali? 2) Dai valori di gNa ricavati, e sapendo che ENa= +55 mV, calcolare i valori corrispondenti di INa e metterli in grafico in funzione di V. (nS) ENa=+55 mV nS gNa=5.6(1-1/[1+EXP((V+30)/6)]) 6.0 5.0 4.0 gNa (nS) 0.00 0.01 0.04 0.19 0.89 2.80 4.71 5.41 5.56 5.59 5.60 5.60 5.60 5.60 5.60 5.60 5.60 Vm -ENa -135 -125 -115 -105 -95 -85 -75 -65 -55 -45 -35 -25 -15 -5 5 15 25 INa (pA) -0.2 -0.9 -4.3 -20.3 -84.5 -238.0 -353.3 -351.5 -305.9 -251.7 -196.0 -140.0 -84.0 -28.0 28.0 84.0 140.0 3.0 2.0 1.0 0.0 -100 -50 0 50 100 50 100 mV pA Vm -80 -70 -60 -50 -40 -30 -20 -10 0 10 20 30 40 50 60 70 80 200 100 0 -100 -50 -100 0 -200 -300 -400 mV Studio dell’inattivazione allo stato stazionario 0 mV 1.2 -60 -80 -100 -120 0 1 2 3 4 Tempo (ms) 5 6 7 0.2 0.0 1.0 0.8 0.6 0.5 0.4 0.2 0.0 -0.2 -120 -0.4 INa (nA) 1 -40 Corrente di Na (nA) Vm (mV) -20 -0.6 -0.8 -1.0 -1.2 0 1 2 3 4 Tempo (ms) 5 6 7 Probabilità gate h aperta 0 0 -100 -80 -60 -40 -20 Potenziale di condiz. (mV) 0 Se si ipotizza che il processo di inattivazione sia regolato da un’unica gate (h), i valori in grafico sono proporzionali alla probabilità di trovare tale gate aperta allo stato stazionario Confronto della voltaggio-dipendenza di gK e gNa 1.0 Conduttanza g 0.8 0.6 0.4 0.2 0.0 -60 -40 -20 0 Vm (mV) 20 40 60 Nota: A parità di conduttanza, è la driving force che determina l'andamento della relazione I/V EA =+55mV Vm -80 -60 -40 -20 0 20 40 60 80 g (nS) 0.00 0.04 0.89 4.71 5.56 5.60 5.60 5.60 5.60 Vm -EA -135 -115 -95 -75 -55 -35 -15 5 25 EB=-80mV IA (pA) -0.2 -4.3 -84.5 -353.3 -305.9 -196.0 -84.0 28.0 140.0 Vm -EB 5 25 45 65 85 105 125 145 165 1000 800 EA=+55mV EB=-80mV Corrente (pA) 600 400 200 0 -200 -400 -100 -80 -60 -40 -20 0 20 Voltaggio (mV) 40 60 80 100 IB (pA) 0.0 0.9 40.0 306.2 472.8 587.9 700.0 812.0 924.0 HHsim: Graphical Hodgkin-Huxley Simulator http://www.cs.cmu.edu/~dst/HHsim/ Esercizio Usando il programma di simulazione H&H, progettare un esperimento di voltage-clamp per determinare: (a) Il potenziale di inversione della corrente del Na (b) Il potenziale di inversione della corrente del potassio Calcolo del potenziale di inversione (equilibrio) della corrente di K+ 80 60 Vtail (mV) IK ist. (nA) Voltaggio (mV) 40 20 0 -20 0 5 10 15 20 25 30 35 -40 V-EK -60 348 -60-EK >0 -70 60 -70-EK >0 -80 -216 -80-EK <0 EK < -60 EK < -70 EK > -80 -60 -80 -100 Tempo (ms) Relazione IK ist. - Voltaggio 400 4000 IK (nA) 3000 2000 1000 0 -1000 0 5 10 15 20 25 30 35 -2000 IK istantanea (nA) 5000 EK 300 200 100 0 -85 -80 -75 -70 -100 -55 -300 Tem po (m s) Voltaggio (mV) IK (nA) -60 -200 -3000 500 400 300 200 100 0 -100 24 -200 -300 -400 -500 -65 +348 nA (-60 mV) +60 nA (-70 mV) 25 26 27 -216 nA (-80 mV) Tempo (ms) 28 29 30 C O
Scarica